Биологические основы вегетативного размножения
Растение представляет собой целостный организм, но в нем отдельные органы, части и ткани различаются между собой по структуре и физиологическим функциям. Присущая растениям способность к регенерации зависит от их внутреннего физиологического состояния и условий внешней среды, однако она проявляется в пределах сорта или клона в неодинаковой степени и может значительно варьировать.
Способность растения восстанавливать утраченные органы или части - наследственный признак, причем способность плодовых и ягодных культур к вегетативному размножению является одним из проявлений их естественной биологической способности к регенерации. Эта естественная регенерационная способность проявляется по- разному, в зависимости от жизненной формы породы (вида, рода). Например, укореняемость полуодревесневших черенков с листьями у деревьев (44,3%), особенно у хвойных и листопадных, ниже. Черенки кустарников укореняются лучше (75,6%), а лианы и травянистые многолетние растения проявляют высокую способность к регенерации придаточных корней (76,8 - 93,1%) (табл. 10).
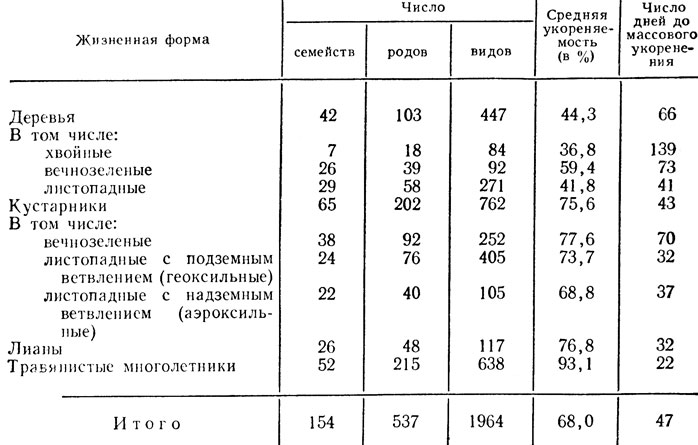
Таблица 10. Укореняемость стеблевых черенков высших растений в зависимости от их жизненной формы (Фаустов, 1977)
Высшие растения характеризуются общностью многих морфологических и физиолого-биохимических признаков, таких, как клеточное строение, дыхание, фотосинтез, направленность основных обменных процессов и т. д. Однако у разных жизненных форм (деревья, кустарники, травянистые многолетники) имеются свои особенности, в значительной мере влияющие на регенерацию и вегетативное размножение. Так, у различных видов и пород наблюдается разная активность эмбриональных тканей в годичном цикле, а в зависимости от жизненной формы - и различная паренхиматизация вегетативных органов, непосредственно влияющая на интенсивность хода восстановительных процессов. В общих чертах можно считать, что чем менее дифференцировано растение, тем активнее и полнее оно восстанавливает свою целостность, и наоборот, высокая специализация и дифференциация коррелируют с меньшей способностью к регенерации. Указанная закономерность проявляется и при индивидуальном развитии особи. Например, растения на ранних фазах онтогенеза проявляют более высокую регенерационную способность, чем на более поздних, а виды и породы, филогенетически более старые, регенерируют труднее, чем филогенетически более молодые, причем у последних способность к восстановлению высокая.
Теоретически каждая соматическая клетка обладает наследственной способностью к восстановлению всего организма, однако потенциальная способность разных культур и их органов к регенерации придаточных органов корней и побегов неодинакова. Замечено, что интенсивность регенерационной способности в значительной степени определяется ростовыми процессами, но на нее оказывают влияние многие внутренние и внешние факторы.
Внутренние факторы являются первостепенными, прямо влияющими на способность изолированных органов или частей растения к восстановлению. У ряда растений (смородина черная, гранат, айва и др.) в стеблевой части побега эндогенно закладываются и затем продолжительное время сохраняют эмбриональную активность группы клеток - меристематические зачатки придаточных корней. У большинства плодовых культур эмбриональные группы клеток возникают также и на корнях, в области перицикла, с дальнейшим формированием придаточных почек, а затем корневых отпрысков.
Меристематические зачатки корней и побегов при поранении растения, нарушении физиологических функций или отделении от материнского растения способны к росту. При этом они соответственно развиваются в придаточные корни или придаточные побеги (корневые отпрыски). На этой способности растения к формированию придаточных органов практически основаны все способы вегетативного корнесобственного размножения. Однако способность к быстрому восстановлению надземной и корневой систем изолированными органами и частями для многих культур зачастую не определяется предварительным эндогенным возникновением меристематических зачатков. Многие древесные и кустарниковые плодовые породы не имеют зачатков, но они довольно быстро их формируют при поранениях, затенении, затоплении или при механическом отделении побегов от растения, то есть при определенных экологических условиях.
Интенсивность новообразования придаточных органов у разных культур разная, она также является наследственным признаком и определяется особенностями формирования вида или породы в филогенезе. По способности к новообразованию придаточных органов все культуры можно условно разбить на три группы:
1) активно образующие придаточные корни и почти не способные к формированию на корнях придаточных побегов (смородина, земляника, крыжовник и др.);
2) интенсивно образующие придаточные побеги на корнях, но трудно формирующие придаточные корни на стеблевых частях (многие сорта яблони, груши, рябина, черешня, орех грецкий и др.);
3) легко восстанавливающие утраченные органы - придаточные корни и побеги (облепиха, лох, айва и др.).
Следует отметить, что среди видов и пород, а внутри вида и сортов существуют большие различия в способности к восстановлению, определяемые не только таксономической принадлежностью, но и внутренним физиологическим состоянием растительного организма.
Способность к регенерации определяется возрастным состоянием особи, онтогенезом побега, взятой для размножения частью или органом и зачастую зависит от направленности обменных процессов в этом органе. Например, высокая укореняемость зеленых черенков большинства косточковых культур совпадает с периодом интенсивного роста побегов в длину, после же формирования побегом верхушечной почки и окончания роста корнеобразовательная способность резко снижается, а после опадения листьев черенки вообще не укореняются. Однако корневые черенки этой же группы культур при оптимальных условиях внешней среды способны практически в течение всего года к придаточному побегообразованию, то есть разные органы растения обладают неодинаковой регенерационной способностью. Листья некоторых пород способны к быстрому укоренению, однако укоренившиеся листья плодовых растений не способны к регенерации побега. Исключение составляют редкие случаи восстановления отделенным листом цитрусовых пород придаточной надземной и корневой систем.
Годичный побег по своей длине проявляет неодинаковую способность к придаточному корнеобразованию, причем различная регенерационная активность зависит от индивидуального развития самого побега, а также от возраста материнского растения. Как правило, растения на ранних этапах своего онтогенеза проявляют высокую регенерационную способность, в дальнейшем она снижается.
Значительное влияние на процессы восстановления придаточных органов оказывает направленность обменных процессов и количественное содержание пластических веществ, особенно углеводов. Имеются убедительные доказательства зависимости физиологического состояния исходного маточного материала и процессов регенерации. Таким образом, регенерация, являясь наследственным признаком, может проявляться по-разному, в зависимости от внутренних факторов роста и развития особи.
Внешние факторы. Способность к вегетативному размножению у плодовых и ягодных культур зависит не только от внутреннего физиологического состояния исходного маточного материала, но и от условий внешней среды (свет, температура, вода, воздушное и минеральное питание, химические регуляторы роста).
Условия освещения оказывают значительное формативное влияние на процессы восстановления. Свет определяет характер развития всех хлорофиллоносных организмов, однако его действие на регенерацию специфично. Так, подземный побег по своему анатомическому строению несколько напоминает корень, вследствие чего такой видоизмененый побег получил название корневища. Этиоляция вызывает накопление природных ауксиновых регуляторов роста в прикамбиальной зоне побега и активизирует появление зачатков придаточных корней. Высокая интенсивность света, наоборот, тормозит дифференциацию эмбриональных клеток в структуры придаточного корня. Однако действие света локальное.
При продолжительном освещении зеленых черенков у них в нижней, затененной зоне формируется больше корней, а в условиях полного затенения корни не развиваются. Вероятно, в данном случае влияние света проявляется в фотосинтетическом накоплении энергопластических веществ, необходимых для закладки придаточных корней. При освещении же корней на них более активно и в большем количестве появляются придаточные побеги. Как известно, ультрафиолетовая часть спектра вызывает разрушение ауксинов, а для закладки и затем дифференциации придаточных почек оптимумом является низкое их содержание в противоположность корневым зачаткам. Приемами этиолирования побегов и даже всего растения пользуются при вегетативном размножении трудно укореняемых культур (яблоня, груша и др.).
Для плодовых и ягодных культур повышенная оводненность растительных тканей и высокая относительная влажность воздуха способствуют протеканию процессов регенерации. Поэтому при искусственных способах вегетативного размножения стремятся к поддержанию оптимального водного режима материнских растений и субстрата. Однако суккулентные растения лучше закладывают придаточные органы при предварительном некотором обезвоживании их в условиях низкой относительной влажности.
По своей направленности обменные физиолого-биохимические процессы при регенерации отличаются от нормально протекающих метаболических реакций в целостном организме. В силу этой специфики для дифференциации эмбриональных клеток и тканей в структуры придаточных органов требуется, как правило, более высокая (на 5 - 7°С) температура. Так, для яблони оптимальной для роста и плодоношения является температура 18 - 24°С, а для укоренения черенков этой породы оптимальная температура колеблется от 24 до 30°С. Однако если зачатки придаточных органов имеются, то оптимумом для их роста является температура 18 - 24°С.
Клебс (1895), а затем Краус и Крейбилл (1918) полагали, что повышенное азотное питание стимулирует вегетативный рост растений, а низкое содержание азота вызывает быструю дифференциацию клеток и их одревеснение с одновременным накоплением в тканях углеводов. В дальнейшем многими исследователями было показано, что избыточное азотное питание тормозит придаточное корнеобразование и почти не влияет на придаточное побегообразование. Повышенное же содержание углекислоты в воздухе эффективно сказывается на процессах регенерации, вероятно, в силу интенсификации фотосинтеза.
Значительное влияние на придаточное органообразование оказывают регуляторы роста, или фитогормоны (ауксины, гиббереллины, цитокинины и ретарданты). Экспериментальными работами многих исследователей была выявлена четкая зависимость новообразования адвентивных корней и побегов от количественного содержания природных ауксинов. В дальнейшем были синтезированы физиологически активные соединения (3-индолилуксусная, индолилмасляная, альфа-нафтилуксусная кислоты и др.), действующие подобно ауксинам. В настоящее время широко исследуется механизм влияния этих регуляторов на процессы регенерации, а сами соединения широко применяются в плодоводстве при вегетативном размножении многих культур.
Интересной особенностью влияния фитогормонов на растения является широкий спектр их действия, зависящий от концентрации. Так, высокие концентрации ауксинов стимулируют новообразование придаточных корней, но тормозят их рост. Для формирования придаточных почек требуется низкая концентрация этого фитогормона, но для их роста - высокая. Гиббереллины и цитокинины во всех концентрациях значительно тормозят дифференциацию придаточных структур, а ретарданты и ингибиторы практически почти не влияют на регенерацию.
Старение и омоложение плодовых растений. Вегетативное размножение в противоположность семенному позволяет в течение многих десятков и сотен лет иметь генетически однородные поколения особей. В плодоводстве их называют сортом, подразумевая под этим термином клон. Клон - совокупность генетически однородных растений, происходящих от одного индивидуума и затем размноженных вегетативно.
Английский ботаник Найт (1816) считал, что клоны при длительном естественном или искусственном вегетативном размножении дегенерируют и вырождаются. Это положение Найта не получило подтверждения. Академик В. Л. Комаров (1940) считает, что "клон не особь и сам по себе не стареет, лишь бы условия жизни его не угнетали". Эта мысль В. Л. Комарова подтверждается существованием многих сортов плодовых деревьев, культивируемых в течение ряда столетий. Многие ученые полагают, что при вегетативном размножении биологическое старение отсутствует в силу омоложения новых особей, причем омоложение особей может быть значительным, вплоть до глубокого, аналогичного омоложению при половом воспроизведении.
Первой теорией, наиболее полно освещающей механизм старения и омоложения при вегетативном размножении высших растений, явилась теория циклического старения и омоложения, разработанная и предложенная независимо друг от друга советскими учеными П. Г. Шиттом (1937) и Н. П. Кренке (1940). Они считают, что при вегетативном размножении каждая часть материнского растения несет возрастные изменения, оказывающие влияние на новое поколение. Однако новые особи начинают свое развитие не с того возрастного этапа, на котором находилось исходное растение или его часть, а с более раннего, то есть они омолаживаются. При анализе причин омоложения в данном случае следует учесть и то обстоятельство, что по своим биологическим свойствам и признакам эмбриональные соматические клетки равноценны половым клеткам. Формирующаяся корневая система оказывает, в свою очередь, омолаживающее влияние на растение.
Таким образом, при длительном вегетативном размножении биологического старения клона не наблюдается, а отдельные особи клона проходят свой цикл индивидуального развития. Одновременно вегетативное размножение позволяет сохранять хозяйственно-ценные особи в ряду поколений, то есть создать клон и прочно закрепить, а затем размножить гетерозиготную форму. Об этом свидетельствует существование многих плодовых растений, например вишни, сливы, инжира, маслины и многих других пород, веками размножаемых вегетативно и сохраняющих до сих пор прекрасные качества плодов.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'