Тепло
Тепловой режим почвенной и воздушной среды является одним из определяющих факторов в обосновании размещения плодовых пород по различным сельскохозяйственным зонам. При этом важное значение имеет не только общее количество тепла, необходимого для нормального роста и развития плодовых растений, но и устойчивость последних к низкой отрицательной (зимостойкость) и высокой положительной (жаровыносливость) температуре. Плодовые породы представляют собой большой и разнообразный набор экотипов, что и определило их неодинаковую требовательность к тепловому режиму.
По признаку теплолюбивости плодовые культуры располагаются следующим образом (в возрастающем порядке): рябина, черемуха, яблоня сибирская, ягодные кустарники (северная зона); земляника, яблоня, вишня, слива, груша (средняя зона); черешня, айва, абрикос, орех грецкий, пекан, фундук, миндаль, персик (южная зона); фисташка, каштан сладкий, хурма, инжир, маслина, цитрусовые (субтропическая зона). Однако следует иметь в виду, что в пределах каждой породы существует большое количество сортов, требования которых к тепловому режиму весьма разнообразны. Так, многие лучшие сорта яблони и груши наиболее успешно произрастают в южной зоне, хотя в целом эти породы более или менее холодостойки.
Отмеченные зоны культивирования плодовых пород соответствуют следующим географическим координатам европейской части СССР: северная зона ограничена 60 - 55° северной широты, средняя зона - 55 - 50°, южная зона - южнее 50° северной широты и субтропическая зона лежит в районах Черноморского побережья Кавказа, Южного берега Крыма и в районах сухих субтропиков Средней Азии. Описанное размещение плодовых пород по зонам следует считать относительно условным.
Так, многие ягодные кустарники успешно возделываются не только в северной, но и в средней зонах. Культура яблони, вишни, сливы распространена как в средней, так и в южной зоне.
Теплолюбивость плодовых пород и сортов определяется двумя факторами: продолжительностью вегетационного периода и режимом температуры. Вполне очевидно, что более теплолюбивые породы и сорта требуют более продолжительного вегетационного периода. Однако при этом важно учитывать режим температуры в течение вегетационного сезона, от которого в значительной мере зависят темпы роста и развития растений.
Так, в Скандинавских странах (Швеция, Дания) на 55 - 60° северной широты вегетационный период достаточно продолжительный и позволяет успешно выращивать многие сорта яблони. Тем не менее целый ряд сортов (Голден Делишес, Джонатан, Старкинг и др.) требует более высокой температуры, чем в Скандинавии, и может успешно выращиваться только в более южных районах.
Одновременно следует учитывать, что плоды многих плодовых культур, в первую очередь яблони и земляники, достигают своих лучших кондиций на высоких широтах. Так, плоды Антоновки обыкновенной приобретают свои лучшие качества при выращивании в Тульской, Рязанской, Тамбовской, Орловской областях, а не в более южных районах. Максимальные урожаи плодов высокого качества сорт Апорт дает в районах с суммой биологически активной температуры (выше 10°С) от 2750 до 3400°С (Белобородова и др., 1973). В то же время продуктивность этого сорта снижается как при избытке (более 3500°С), так и при недостатке (менее 2700°С) тепла.
Продолжительность вегетационного периода и температурный режим оказывают влияние не только на продуктивность плодовых культур и качество урожая. От этих факторов в значительной мере зависит устойчивость растений к неблагоприятным условиям среды. Так, при недостатке тепла плодовые растения не успевают закончить свое развитие, древесина их не вызревает, в результате чего они не могут полностью пройти этапы осеннего и раннезимнего закаливания к отрицательной температуре.
Требования плодовых культур к температурному режиму неодинаковы в течение вегетационного периода. Они наиболее чувствительны к теплу в период цветения и в первые дни после начала развития плодов. Это связано прежде всего с процессом оплодотворения и формированием зародыша. Так, при температуре 21°С прорастание пыльцевой трубки в столбике пестика у сорта груши Анжоу завершается в течение 24 ч, а при температуре 16,5 и 10°С - в течение 72 и 120 ч соответственно (Mellenthin et al., 1972). Среднедневная температура ниже 14 - 15°С в первые дни после цветения существенно уменьшает количество семян в плодах этого сорта и в 1,5 - 2 раза снижает подуктивность деревьев. Общеизвестна повышенная требовательность к тепловому режиму в период цветения и других плодовых пород.
Высокая потребность плодовых растений в тепле отмечается в период закладки цветковых почек. Установлено, что для яблони в этот период необходима среднесуточная температура не ниже 18-20°С (Коломиец, 1966). Высокая температура в этот период определяет не только начало закладки генеративных органов, но и способствует формированию большого количества цветков в почке. Кроме того, у яблоки, например, более высокая температура вызывает закладку цветков в пазушных почках побега.
Следовательно, у плодовых пород отмечается по крайней мере два критических периода в их отношении к теплу. Оба они связаны с развитием генеративных органов, причем в первом случае критический период определяется оплодотворением и развитием зародыша, во втором связан с закладкой цветковых почек.
На рост и развитие плодовых растений и их продуктивность отрицательное влияние оказывает и избыток тепла. Температура выше 30 - 35°С угнетающе действует на рост и развитие многих плодовых культур, сложившихся в условиях умеренно теплого климата. Более высокая температура(выше 50°С) приводит к повреждению коры дерева и ожогу плодов, особенно у крупноплодных сортов яблони и груши.
Реакция плодовых культур на высокую температуру определяется их жароустойчивостью. У менее устойчивых пород и сортов жаркая погода вызывает распад белков протоплазмы, нарушает белково-липоидный комплекс и субмикроскопическую структуру протопласта. Все это может привести к отмиранию не только тканей и отдельных органов, но и к гибели всего растения (Генкель, 1967).
Продолжительное воздействие избытка тепла на плодовые растения может способствовать неравномерному росту плодов и неодновременному их созреванию, ухудшает их покровную окраску, снижает вкусовые качества и уменьшает лежкость. Под влиянием высокой температуры зачастую увеличивается доуборочное опадение плодов и поражаемость плодовых растений некоторыми вредителями и болезнями.
Потребность плодовых растений в пониженной температуре. Филогенез основных плодовых пород, возделываемых на территории СССР, протекал в умеренных широтах, которым свойственны определенные закономерности в колебаниях теплового режима погоды в годичном цикле. В этих условиях у плодовых растений наследственно закрепился соответствующий ритм роста и развития. Важным свойством этого ритма является состояние так называемого покоя, в сущности представляющего собой приспособительную реакцию растений к неблагоприятным факторам среды и в первую очередь к перезимовке.
Нормальный ход процессов роста и развития у плодовых растений возможен только при обязательном воздействии пониженной температурой (около 0°С). Это воздействие необходимо для прерывания состояния покоя, после чего растения приобретают способность к нормальному росту и развитию на всем протяжении вегетационного периода.
Без воздействия пониженной температурой плодовые растения длительное время остаются в состоянии глубокого органического покоя, то есть не трогаются в рост при наличии необходимых благоприятных условий. Так, деревья яблони, не подвергнутые воздействию пониженной температурой, не трогаются в рост (остаются в состоянии покоя) в течение 140 - 200 дней. Кроме этого, у них наблюдается растянутый период цветения (от февраля до сентября) и отмечается израстание соцветий (Поплавская, Голубкова, 1961).
Недостаточное охлаждение является серьезной помехой при выращивании персика в районах с теплыми зимами (США, Австралия). Так, в годы, когда сумма часов с температурой ниже 7,2°С за декабрь и январь не превышает 400, наблюдаются аномалии в развитии цветковых почек, в результате чего большая часть их опадает и деревья остаются без урожая.
Обработка плодовых растений пониженной температурой широко применяется в практике. В первую очередь это относится к выгоночной культуре земляники в защищенном грунте, выращивание которой практически невозможно без выдерживания растений в течение определенного времени при пониженной температуре. Сюда же можно отнести и специальную обработку (стратификация) семян плодовых растений пониженной температурой.
Продолжительность периода глубокого покоя, а следовательно и продолжительность воздействия пониженной температурой, неодинакова у плодовых культур. Обычно она связана с экологическими условиями, в которых проходило развитие той или другой плодовой породы или формы. Так, для пород и сортов восточных экотипов период пониженной температуры значительно короче, чем для более западных и южных экотипов. Это объясняется тем, что в восточных районах осенний период до наступления устойчивых морозов менее продолжительный. Следовательно, в этих условиях растения находятся меньше времени при пониженной положительной температуре, чем в западных и южных районах. Эта особенность эволюционно явилась, по-видимому, основным фактором в выработке растениями умеренных широт глубины и продолжительности покоя.
Физиологическая сущность воздействия пониженной температурой на плодовые и другие многолетние растения, вероятно, имеет много общего с природой озимости у зерновых культур. Очевидно, это связано прежде всего с деятельностью ферментной системы растения, направленность действия которой может изменяться под влиянием температуры окружающей среды; под влиянием пониженной температуры происходит усиление гидролитических и окислительных процессов, мобилизация и преобразование запасных веществ, накопление энергии в виде АТФ. Важным моментом является смещение в балансе регуляторной системы в сторону усиления функций стимуляторов роста (гиббереллины, ауксины, цитокинины) и снижения функций ингибиторов роста (абсцизовая кислота).
Зимостойкость и морозоустойчивость плодовых растений. Зимние повреждения плодовых растений встречаются повсеместно в районах промышленного плодоводства. В отдельные годы они принимают массовый характер и приносят большой ущерб народному хозяйству. Установлено, что отрицательная температура наносит сильный вред плодовым растениям в зимний период один раз в 6 - 8 лет. Кроме того, в отдельных районах ежегодно наблюдаются менее значительные повреждения, прежде всего подмерзание цветковых почек у косточковых культур в зимний период и подмерзание цветков во время весенних заморозков, также приносящие заметный ущерб плодоводству. Поэтому проблема зимостойкости плодовых культур в нашей стране имеет важное народнохозяйственное значение.
Зимостойкость - понятие сложное, оно характеризует устойчивость плодовых растений не только к отрицательной температуре, но и ко всему комплексу погодных условий в течение зимы. Общая устойчивость к зимним повреждениям определяется не только морозоустойчивостью, то есть потенциальной (наследственной) устойчивостью породы или сорта к отрицательной температуре, но и соответствием ритма их роста и развития ритму изменения погодных условий в годичном цикле.
Наиболее полно потенциальная морозоустойчивость и в целом зимостойкость проявляются в конкретных условиях среды, в которой протекало формирование породы или сорта. При выращивании плодовых растений в других условиях среды, не соответствующих ритму их развития, они могут в значительной мере утратить свою зимостойкость. Так, потенциально морозоустойчивые сорта и породы северовосточных экотипов могут быть слабозимостойкими в средней зоне европейской части СССР, где зима значительно мягче, но ритм колебания погодных условий в течение года существенно отличается.
Степная вишня достаточно морозоустойчива в Поволжье и в районах Урала. Однако в средней зоне у нее часто подмерзают цветковые почки, особенно после зимних оттепелей. Яблоня Антоновка обыкновенная - высокозимостойкий сорт в средней полосе СССР, а на Кубани повреждается заморозками. Подобных примеров можно привести немало. Следовательно, зимостойкость надо рассматривать как взаимодействие растительного организма с факторами окружающей среды.
Устойчивость плодовых растений к отрицательной температуре в значительной мере зависит от температурного и светового режимов, от количества осадков и их распределения в течение вегетационного периода. Этими факторами определяется не только общее развитие плодовых растений, но и своевременное прекращение их роста, что необходимо для вызревания древесины и прохождения процессов закаливания, обусловливающих их подготовку к перезимовке.
Механизм морозоустойчивости плодовых растений. Сложность физиолого-биохимической сущности зимостойкости растений обусловливает большие трудности в разработке общей биологической теории механизма устойчивости растений к отрицательной температуре. Однако достижения науки последних лет позволяют в значительной мере познать природу морозоустойчивости и факторы, определяющие ее.
Одной из главных причин губительного действия отрицательной температуры является образование внутриклетного кристаллического льда. Этот процесс может иметь место или при очень быстром охлаждении, или при длительном воздействии низкой отрицательной температурой. При быстром понижении температуры вода не успевает проникнуть в межклетники и лед образуется между протопластом и клеточной оболочкой. В этом случае гибель клетки наступает при оттаивании в результате повреждения плазмалеммы и потери ею проницаемости. При очень резком падении температуры кристаллы льда возникают в самой протоплазме и вызывают ее разрывы и гибель наступает уже во время замораживания (Бугаевский, 1955; Самыгин, 1960, 1964).
В природных условиях губительное действие быстрого замораживания чаще всего наблюдается в весеннее время или после зимних оттепелей. Наиболее опасные повреждения возникают при длительном воздействии низкой отрицательной температурой. В таких условиях, когда часть оставшейся воды гораздо медленнее перемещается в межклетники, появляется опасность образования кристаллов льда внутри клетки. По мнению О. А. Красавцева (1974), при длительном охлаждении гибель клетки происходит не из-за ее обезвоживания, на что иногда указывают некоторые исследователи, а в результате образования очень мелких кристаллов внутриклетного льда, которые повреждают субмикроскопические элементы клетки и оказывают отрицательное действие на метаболические процессы. Работы, выполненные в лаборатории И. И. Туманова, показывают, что повреждающее действие при медленном охлаждении может оказывать кристаллический лед, образующийся и в межклетниках, вызывая деформацию клеточных стенок.
Губительное действие кристаллического льда убедительно подтверждается исследованиями по витрификации живых организмов. Показано, что в соответствующих условиях, препятствующих кристаллизации льда, растения могут выдерживать очень низкую отрицательную температуру. Так, ветви древесных растений, в том числе и плодовых, после закаливания и охлаждения до соответствующей температуры могут выживать после погружения их в жидкий азот и другие жидкости, имеющие температуру от - 195 до - 250°С (Туманов и др., 1959).
В процессе эволюции у растений выработались защитные свойства против губительного действия отрицательной температуры. Н. А. Максимов (1913) впервые обосновал положение о химической защите клеток растворами Сахаров и некоторых солей, снижающих эвтектическую точку, то есть температуру замерзания раствора. В настоящее время установлено, что метаболизм углеводов занимает важное место в механизме морозоустойчивости растений. Однако следует иметь в виду, что сахара непосредственно не определяют устойчивость растений к отрицательной температуре. Защитная роль их в полной мере определяется только при соответствующем физиологическом состоянии растений, которое приобретается в процессе закаливания. Этим, очевидно, объясняются факты, когда количественное содержание Сахаров не является достаточно надежным критерием при оценке зимостойкости разных сортов яблони (Тучков, 1970).
В последние годы защитную роль Сахаров рассматривают не только в связи со снижением эвтектической точки. Большое значение придается роли Сахаров в повышении устойчивости белков к денатурации при воздействии отрицательной температурой (Хеббер, 1964). Считается, что сахара повышают устойчивость ферментных систем, участвующих в процессах фосфорилирования. Большого внимания заслуживает также предположение У. Хеббера о том, что защитное действие Сахаров проявляется только в том случае, если они находятся в непосредственном контакте с белками, хотя сам механизм этого действия остается еще не выясненным.
В механизме морозоустойчивости растений определенная роль принадлежит полимерным углеводам, в первую очередь гемицеллюлозам.
Установлено, что внутренние слои клеточных стенок увеличиваются к осени и уменьшаются к весне (Parkez, 1963). Это дает основание предполагать, с одной стороны, участие в этом процессе гемицеллюлоз, а с другой,- положительное значение этого явления в механизме морозоустойчивости растений в результате стабилизации клеточных оболочек.
В настоящее время выявлена зависимость морозоустойчивости от фенольных соединений, в частности от содержания флоридзина (Сарапуу, Томм, 1970). Роль последнего, по-видимому, связана с его участием в образовании лигнина, способствующего одревеснению и вызреванию тканей, что необходимо для закаливания растений.
Важная роль в механизме морозоустойчивости принадлежит белкам, являющимся главной жизненной субстанцией живой клетки. Исследованиями У. Хеббера (1952, 1964) установлено, что не все белки обладают одинаковой устойчивостью к отрицательной температуре. Это позволило сформулировать денатурационную теорию морозоустойчивости. При этом высказывается предположение, что механизм повреждения клеток определяется количеством белка с сульфгидрильными группами и их устойчивостью к окислению, то есть к образованию межмолекулярных дисульфидных связей, что может привести к развертыванию белковых молекул и их денатурации (Левитт, 1966).
В настоящее время гипотеза денатурации белков позволяет считать, что устойчивость протоплазмы есть функция резистентности протеинов (Браун и др., 1964). Однако эта гипотеза объясняет механизм морозоустойчивости односторонне, основываясь только на чувствительности белков к отрицательной температуре. Очевидно, важная роль белков в механизме устойчивости проявляется в их функциональной деятельности в цитоплазме в процессе закаливания растений к низкой температуре. В этой связи представляет интерес и тот факт, что увеличение количества белков с сульфгидрильными группами может привести к усилению гидролитических процессов в растениях, протекающих в период закаливания, поскольку SH-группы входят в состав таких ферментов, как амилаза, фосфорилаза и др.
В последние годы большое значение в механизме морозоустойчивости придается липидам, входящим в состав клеточных мембран. По мнению Дж. Левитта (1966), нарушение липопротеиновых комплексов мембран цитоплазмы в процессе замерзания приводит к более быстрой гибели клетки, чем в случае денатурации белков.
Физико-химические свойства мембран во многом зависят от состава жирных кислот, входящих в состав липидов (липопротеинового комплекса мембран). В ходе подготовки растений к перезимовке значительно возрастает количество ненасыщенных жирных кислот и повышается их отношение к насыщенным (Ketehil, 1966). Поскольку температура плавления ненасыщенных жирных кислот ниже, чем насыщенных, можно полагать, что повышение содержания первых способствует стабилизации проницаемости мембран, увеличивает эластичность протоплазмы и этим, вероятно, определяет устойчивость клетки к повреждению при замерзании.
В механизме морозоустойчивости растений важное значение имеют нуклеиновые кислоты. Их роль определяется, с одной стороны, непосредственным участием в синтезе белков, от чего зависит качественный состав последних, а с другой,- непосредственной устойчивостью молекул нуклеиновых кислот к отрицательной температуре. Поскольку молекула ДНК более устойчива к замерзанию, чем молекула РНК (Shikama et al., 1965), это дает основание считать, что низкая отрицательная температура в большей мере нарушает синтез РНК и белка, чем синтез ДНК.
Устойчивость растений связывают с изменением качественного состава РНК. Высокое содержание гуанина и цитозина в молекуле РНК, а также способность растений синтезировать РНК с высоким содержанием этих нуклеотидов занимает важное место в процессе подготовки растений к засухо- и холодоустойчивому состоянию. Однако преобладание в молекуле РНК отмеченных нуклеотидов как фактора стабильности РНК, определяющего морозоустойчивость растений, вероятно, следует рассматривать в пределах вида или даже сорта. Что касается различий между видами, то эти закономерности не подтверждаются (Белозерский, Спирин, 1960). Возможно, морозоустойчивость растений в большей мере связана не вообще с особенностями нуклеотидного состава РНК, а с последующим его изменением в процессе закаливания.
Физико-химические свойства клеток, определяющие их морозоустойчивость, во многом зависят от содержания воды и ее связи с молекул ярно-структурными элементами протоплазмы и в целом клетки. В процессе подготовки растений к перезимовке повышение устойчивости к отрицательной температуре положительно коррелирует с возрастающей водоудерживающей способностью тканей. Это можно объяснить накоплением гидрофильных коллоидов в протоплазме, главным образом пентозанов.
Однако данные о связи устойчивости разных сортов с их водоудерживающей способностью противоречивы. Наряду с положительной зависимостью в ряде случаев приводятся сведения, в которых эта связь не подтверждается. Представляет интерес и тот факт, что к весне у плодовых деревьев водоудерживающая способность увеличивается, а морозоустойчивость существенно снижается. Отмеченное противоречие можно объяснить тем, что применяемые до сих пор в большинстве случаев методы определения водоудерживающей способности тканей позволяют судить не столько об истинной прочности связи воды с молекулярными и структурными элементами клетки, сколько вообще об оводненности тканей. Последний же показатель довольно непостоянен и зависит от условий окружающей среды. Кроме этого, водоудерживающая способность во многом определяется и морфолого-анатомическими особенностями побегов, в частности развитием их покровных тканей. Так, слабозимостойкие сорта сливы Персиковая и Ренклод Альтана с хорошо развитыми покровными тканями теряют при подсушивании меньше воды, чем более устойчивые сорта (Максименко, 1971).
Во многих работах последних лет достаточно убедительно показано, что с повышением морозоустойчивости в тканях растений увеличивается количество прочно связанной воды. Это происходит в результате молекулярно-структурных изменений клетки и прежде всего ее протоплазмы (Сулейманов, 1964). Физико-химические изменения в клетках, происходящие под влиянием понижающейся температуры, способствуют переходу воды в новое структурное состояние, когда уменьшается ее поступление на образование межклетного и внутриклетного льда при замерзании (Mazur, 1965).
В другом аспекте рассматривается зимостойкость растений в связи с так называемой зимней засухой. В этом случае гибель растений происходит вследствие их чрезмерного иссушения в зимний и ранне- весенний периоды, когда при замерзшей почве устанавливается солнечная погода и температура днем поднимается выше 0°С, что приводит к увеличению транспирации. Наиболее часто зимне-весеннее иссушение плодовых деревьев наблюдается в южных районах, особенно с холмистым рельефом местности.
В настоящее время механизм иссушения растений еще недостаточно выяснен. Однако исследования Л. К. Константинова и В. С. Шкреба (1971) дают основание считать, что иссушение отдельных участков дерева (ствола, оснований скелетных ветвей) происходит не только в результате испарения, но и вследствие перераспределения воды из-за градиента температуры на разных участках скелетной ветви.
Повышение устойчивости растений к отрицательной температуре.
Морозоустойчивость растений - сложное физиологическое свойство. Исследования И. И. Туманова (1940) и его школы показали, что это свойство у растений развивается в течение всей вегетации, но особенно сильно в позднелетний и осенний периоды, на основании чего была разработана теория закаливания к отрицательной температуре, получившая всеобщее признание.
Основные положения теории И. И. Туманова указывают на то, что развитие морозоустойчивости у высших растений проходит поэтапно. Первый этап характеризуется ослаблением и прекращением РОСТОВЫХ процессов и вхождением растений в состояние покоя. Только при этом условии возможно дальнейшее повышение устойчивости растений к отрицательной температуре.
Ритмичность погодных условий в годичном цикле оказывает большое влияние на характер перехода растений в состояние покоя, прежде всего на его так называемую глубину и продолжительность. Развитие этих свойств явилось реакцией растений на изменения условий среды, что, возможно, лежит в основе природы адаптации их к неблагоприятным факторам, в том числе и к отрицательной температуре.
Следующий этап развития морозоустойчивости И. И. Туманов называет первой фазой закаливания, которая в средней полосе протекает в первой половине осени (сентябрь, октябрь). Решающее влияние на закаливание в этот период оказывает постепенное понижение температуры до низкой положительной и хорошее освещение, благоприятствующее фотосинтезу. Именно в этот период в основном начинается соответствующая перестройка субмикроскопических структурных элементов и высокомолекулярных соединений клетки и происходит накопление защитных веществ.
Для дальнейшего повышения морозоустойчивости необходима вторая фаза закаливания (третий этап развития морозоустойчивости), которая протекает в условиях постепенного понижения отрицательной температуры. Плодовые деревья в средней полосе СССР проходят вторую фазу обычно в течение ноября. Эта фаза у древесных растений является наиболее эффективной.
Морозоустойчивость плодовых деревьев не остается постоянной в течение зимнего периода. Обычно она выше в начале зимы (декабрь) и снижается к весне, хотя это наблюдается не всегда. Так, в средней полосе СССР устойчивость яблони в зиму 1968/69 г. в марте была выше, чем в феврале (Агафонов, Кладько, 1974). В данном случае повышению морозоустойчивости способствовала продолжительная и довольно устойчивая погода с очень низкой отрицательной температурой в предшествующие месяцы (январь, февраль). Как показывают исследования И. И. Туманова и О. А. Красавцева (1959), по мере снижения температуры после второй фазы закаливания устойчивость растений к отрицательной температуре повышается.
В зиму 1970/71 г. морозоустойчивость яблони в феврале по сравнению с декабрем существенно увеличивалась. При этом повышение устойчивости отмечалось после продолжительной оттепели (две декады января), когда имело место раззакаливание растений. Однако в результате постепенного и продолжительного понижения температуры после оттепели возобновилась и продолжилась вторая фаза закаливания, что и обеспечило не только восстановление морозоустойчивого состояния, но и способствовало его повышению. Важно обратить внимание на тот факт, что повторное закаливание проходило в то время, когда деревья яблони вышли из состояния глубокого органического покоя. Следовательно, для развития морозостойкого состояния у древесных растений более решающее значение имеет не глубокий органический покой, а покой вообще.
Особенности подмерзания надземной части у плодовых деревьев. В основном наблюдаются три вида повреждений плодовых деревьев отрицательной температурой: подмерзание обрастающих ветвей (периферии кроны), штамба и основания скелетных ветвей и подмерзание цветков и завязей в период весенних заморозков. Наиболее губительное из них - подмерзание штамба и скелетных ветвей.
Подмерзание обрастающих ветвей встречается наиболее часто. Можно считать, что в той или иной мере побеги повреждаются морозами практически ежегодно. Чаще всего эти повреждения затрагивают ткани древесины, реже ткани коры и гораздо реже клетки камбия. Подмерзшие клетки и ткани обычно приобретают бурую или коричневую окраску и хорошо заметны на поперечных и продольных срезах.
При слабом подмерзании обрастающих ветвей, когда остаются живые клетки коры, камбия и почти не нарушается проводящая система, плодовые деревья быстро восстанавливают поврежденные ткани и практически не страдают после перезимовки. Однако в суровые зимы повреждения бывают значительными, что приводит к усыханию прежде всего старых плодоносных ветвей (плодух), побегов и даже более крупных обрастающих ветвей. Нередко после сильного подмерзания ветви остаются живыми, но рост и развитие их в последующие годы бывают значительно ослабленными, и часто такие ветви отмирают на 2 - 3-й год после подмерзания. Обычно это связано с повреждением (закупоркой) проводящей системы камедеобразными веществами. У косточковых более часто на обрастающих ветвях кроны подмерзают цветковые почки.
Повреждения штамба и основания скелетных ветвей бывают в основном двух видов. В одном случае эти части дерева подмерзают в начале и середине зимы, в другом повреждения отмечаются ранней весной и в плодоводстве называются солнечными ожогами. В обоих случаях повреждения затрагивают ткани коры и камбия. Это приводит к омертвению значительных участков штамба и скелетных ветвей или к усыханию всего дерева по уровень снегового покрова. Нередко после таких повреждений у плодовых деревьев отмирают отдельные скелетные ветви.
Подмерзание в начале и середине зимы связано главным образом с плохим вызреванием тканей. Исследования М. А. Соловьевой (1967) показывают, что активная деятельность клеток камбия у основания скелетных ветвей, особенно в развилках, заканчивается позже по сравнению с другими частями дерева. В результате эти части дерева не успевают в полной мере пройти процесс закаливания и не приобретают высокой морозоустойчивости. То же самое можно отнести и к тканям нижней части штамба и зоны корневой шейки. Отмеченные повреждения чаще всего затрагивают молодые плодовые деревья.
Нередко повреждения штамба и оснований скелетных ветвей проявляются в виде морозобоин, представляющих собой глубокие трещины (разрывы коры и древесины), доходящие иногда до центра дерева. Такие повреждения чаще всего бывают при сильных морозах, особенно при резких колебаниях температуры днем и ночью. Очевидно, растрескивание древесины и коры является следствием интенсивного образования большого количества кристаллического льда в тканях дерева. Причина этих повреждений, по-видимому, также лежит в факторах, препятствующих прохождению плодовыми деревьями процесса закаливания. Чаще всего их связывают с затяжным характером ростовых процессов.
Ожоги штамба и основания скелетных ветвей наиболее часто встречаются в континентальных районах. При легких повреждениях погибают только наружные ткани коры, при более тяжелых отмирают целиком вся кора, камбий и внешние слои древесины. Ожоги проявляются в виде омертвевших участков с южной и юго-западной сторон дерева, иногда захватывающих более половины ствола по окружности.
Солнечные ожоги вызываются резкими колебаниями температуры днем и ночью, которые довольно часто наблюдаются в конце зимы и начале весны. В средней полосе СССР резкие перепады температуры обычно отмечаются в марте. В ясную солнечную погоду температура коры крупных ветвей и штамба с южной стороны может подниматься до 15 - 20°С. После захода солнца температура резко падает и может понижаться ночью до - 20°С. В результате перепад температуры в течение суток достигает 40°С и более.
Причину солнечных ожогов чаще всего видят в том, что под влиянием нагрева происходит локализованный выход тканей коры и камбия из состояния покоя. В результате этого они утрачивают морозостойкое состояние и повреждаются при значительном понижении температуры ночью (Соловьева, 1967). Однако эти доводы недостаточны для более полного объяснения причин, вызывающих ожоги у плодовых деревьев. Исследования, проведенные в последние годы, дают основание полагать, что эти повреждения могут вызываться и иссушением тканей, происходящим в результате испарения и перераспределения воды из-за градиента температуры на разных участках крупных ветвей (Константинов, Шкреба, 1971).
Особенности подмерзания корневой системы. По сравнению с надземной частью дерева корни обладают значительно меньшей морозоустойчивостью. Так, при благоприятных условиях закаливания ветви плодовых пород, произрастающих в средней полосе СССР, могут переносить морозы до - 35 - 40°С, а корни, как правило, выдерживают не более - 15 -16°С. Однако необходимо помнить о том, что при соответствующих условиях закаливания корни приобретают такую же устойчивость к отрицательной температуре, как и надземная система дерева (Туманов, Хвалин, 1967).
Корневая система повреждается морозами значительно реже, чем надземная часть дерева. Наиболее существенные подмерзания наблюдаются в суровые и малоснежные зимы, особенно в первую половину зимы, когда корни еще не прошли в полной мере вторую фазу закаливания. При этом более заметные повреждения отмечаются после сухой второй половины лета и осени, хотя причины этого еще не установлены.
Повреждения корней морозами бывают разные. В одних случаях подмерзают ткани древесины, а клетки коры и камбия остаются живыми. Как правило, после таких повреждений корни восстанавливаются. Однако в зависимости от степени подмерзания дерево может в той или иной мере страдать. Проявляется это в более позднем распускании почек, ослаблении ростовых процессов, осыпании цветков и завязей.
Наиболее опасны повреждения, затрагивающие клетки коры и камбия. В этом случае омертвевшие участки коры отстают от древесины, корни оголяются и отмирают. После значительного подмерзания корневая система, как правило, не восстанавливается и дерево в первый же год после суровой зимы погибает.
Повреждение цветков и завязей весенними заморозками. Поздние весенние заморозки наносят большой ущерб промышленному плодоводству. Их действие в основном проявляется в период цветения плодовых культур или в период формирования завязей. В это время заморозки могут привести к полной гибели урожая или вызвать существенные дефекты в развивающихся плодах вследствие частичного подмерзания их тканей.
Наиболее чувствительны к заморозку пестики и семяпочки. В связи с этим по внешнему виду иногда бывает трудно определить поврежденные цветки, поскольку лепестки и тычинки не подмерзают и сохраняют нормальный вид даже после значительных заморозков. Однако при внимательном наблюдении можно заметить поврежденные органы: пестик имеет побуревший вид, а на разрезе завязи видны потемневшие семяпочки, в результате чего в таких цветках не происходит оплодотворение и они отмирают. Только у пород и сортов плодовых культур, склонных к партенокарпии, возможно развитие бессемянных плодов после заморозков.
Нередко заморозки повреждают завязи и молодые развивающиеся плоды. В этом случае плоды могут не опадать. Однако частичное повреждение тканей плода чаще всего приводит к образованию в верхней части его участков опробковевшей кожицы. Иногда появляются вздутия и трещины, в результате чего плоды в значительной мере утрачивают свои товарные качества.
Несмотря на довольно заметные различия в морозоустойчивости между отдельными плодовыми породами, критические отрицательные температуры для цветков и завязей во время заморозков лежат в близких пределах (табл. 8). Это, очевидно, связано с тем, что в период цветения плодовые деревья вышли из морозостойкого состояния (произошло раззакаливание) и различия в морозоустойчивости между ними сглаживаются.
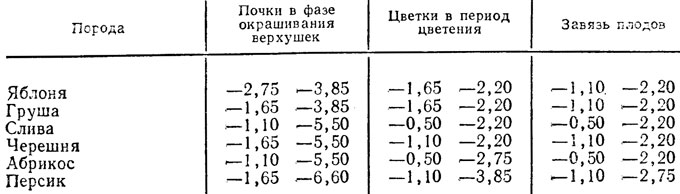
Таблица 8. Критические температуры (в °С) во время распускания цветковых почек и цветения различных плодовых пород
Пути повышения морозоустойчивости плодовых растений. Изучение механизма морозоустойчивости открывает возможности направленного воздействия на растения с целью регулирования генотипического проявления этого свойства. Морозоустойчивость определяется прежде всего наследственно-биологическими особенностями. Поэтому в решении проблемы зимостойкости в растениеводстве главное внимание следует уделять как подбору соответствующих сортов, имеющихся в производстве, так и созданию новых форм, более устойчивых к неблагоприятным факторам зимнего периода.
Проявление наследственных признаков, обусловливающих потенциальную устойчивость растительного организма к отрицательной температуре, в значительной мере зависит от условий окружающей среды, в" том числе и от агротехники. Содержание почвы в междурядьях сада, система удобрений, орошение, обрезка, регулирование плодоношения, тип почвы оказывают существенное влияние на морозоустойчивость.
Значительное влияние на термический режим и микроклимат в целом оказывает рельеф местности, что в немалой степени определяет характер зимних повреждений. Продолжаются исследования по изучению роли удобрений, в том числе отдельных элементов. Подтверждено влияние калия (особенно при сочетании калийных и полного удобрений) на повышение морозоустойчивости (Соловьева, Оканенко, 1968). В настоящее время большое внимание уделяют изучению роли микроэлементов в морозоустойчивости растений, особенно магния и бария.
Важное значение имеет создание условий, способствующих более быстрому преодолению последствий зимних повреждений: поддержание оптимального режима влажности почвы в течение вегетации, обрезка.
После зимних повреждений рекомендуется осторожно использовать гербициды, так как контакт их с подмерзшими частями может препятствовать развитию молодых восстанавливающихся тканей.
Важное значение для зимостойкости земляники имеют сроки посадки. Поздние сроки посадки (позже августа в средней полосе СССР) препятствуют хорошему развитию растений до наступления морозов. В суровые зимы, особенно при недостаточном снеговом покрове, это является одной из причин значительного вымерзания растений.
В зимостойкости плодовых растений значительную роль играет подвой. В настоящее время известно, что подвой нередко оказывает существенное влияние на зимостойкость привоя. Так, исследования Н. В. Субботиной и В. Я. Браду (1969) показали, что у деревьев персика, привитых на алыче, зимостойкость цветковых почек снижается в большей степени, чем у привитых на сеянцы абрикоса. Подвой воздействует на изменение морозоустойчивости и у деревьев яблони. Так, у сортов Голден Делишес и Старкинг, привитых на MVII, отмечалось значительное подмерзание тканей флоэмы, а у привитых на сеянцевые подвои повреждений не наблюдалось (Simons, 1970).
В зимостойкости косточковых пород важное значение имеет корнесобственная культура. Кор несобственные растения способны довольно быстро восстанавливаться после подмерзания за счет формирования придаточных почек и затем отрастания корневой поросли.
Для зимостойкости привитых плодовых деревьев важное значение имеет и устойчивость к отрицательной температуре самих подвоев и в первую очередь их корневой системы. Одной из причин недостаточной морозоустойчивости сеянцев зимостойких сортов является то, что в результате значительной генетической разнокачественности семенная популяция подвоев может обладать существенной амплитудой колебания по признаку морозоустойчивости. В настоящее время в насаждениях яблони зимостойкие сорта, семена которых используют для выращивания подвоев, нередко опыляются слабозимостойкими (например, совместное выращивание Антоновки и Пепина шафранного). Роль опылителя в повышении морозоустойчивости семенных подвоев значительна. Особенно это наглядно проявилось в суровую и малоснежную зиму 1968/69 г., когда в результате подмерзания корневой системы сильно пострадали сады в средней полосе страны.
В плодоводстве большое внимание уделяют повышению морозоустойчивости клоновых подвоев. Наряду с успешной селекцией на зимостойкость с этой же целью начинают применять выращивание плодовых деревьев с промежуточной вставкой клонового подвоя. Однако удовлетворительные результаты получают лишь в том случае, если вставка клонового подвоя размещена ниже уровня почвы, что предохраняет ее от вымерзания (Шляпников, 1970).
Подмерзание штамба и оснований скелетных ветвей чаще наблюдается у сортов, не обладающих достаточной устойчивостью к низкой отрицательной температуре. Опыт показывает, что прививка слабозимостойких сортов на штамб или в крону зимостойкого скелетообразователя может повысить их устойчивость к повреждающему действию отрицательной температуры.
В настоящее время широкие эксперименты по культуре яблони на скелетообразователях проводятся на кафедре плодоводства ТСХА (Анзин, Сусов, 1971, 1972). Результаты опытов показывают, что с помощью прививки на зимостойкие скелетообразователи (Антоновка обыкновенная, Коричное полосатое, Шаропай, некоторые формы китаек) появляется реальная возможность выращивать в средней полосе СССР целый ряд слабозимостойких сортов яблони (Уэлси, Мелба, Лобо, Спартан, Пепин шафранный и др.), обладающих хозяйственно-ценными признаками.
В США с целью защиты штамба и скелета деревьев персика начинают разрабатывать приемы с использованием специальных теплоизоляционных материалов. В частности, появляется возможность защиты от солнечных ожогов с помощью полиуретана, покрытого снаружи специальным отражателем, и алюминиевой фольги со стекловолокном (Sawage, 1970).
Существующая зависимость между процессами роста и морозоустойчивостью открывает возможность использования физиологически активных веществ, позволяющих направленно влиять на рост и развитие растений и таким путем изменять их устойчивость к отрицательной температуре. Применяя физиологически активные вещества (регуляторы роста), ставят следующие цели:
тормозя ростовые процессы (использование ретардантов), повысить морозоустойчивость растений с продолжительным периодом роста;
замедляя темпы развития частей цветка, повысить морозоустойчивость цветковых почек у косточковых культур;
проводя химическое прореживание цветков и завязей при обильном цветении, повысить устойчивость всего дерева;
путем ускорения оттока пластических веществ из листьев в зимующие органы увеличить в последних содержание защитных веществ.
В настоящее время известно, что опрыскивание молодых деревьев груши 1%-ным раствором хлорхолинхлорида наряду с торможением роста способствует повышению их морозоустойчивости (Modlibowskaj, 1961).
Положительное влияние на повышение морозоустойчивости оказывает и другой ретардант - алар. В экспериментах Научно-исследовательского зонального института садоводства нечерноземной полосы показано, что обработка аларом саженцев в питомнике сокращает период их роста и способствует повышению морозоустойчивости коры и почек (Егураздова, Тюрина, 1971). В опытах с малиной также установлено положительное влияние ретардантов на повышение морозоустойчивости. Так, обработка растений в начале последней декады июня или в середине июля аларом в дозе 2000 мг/л позволяет в 2 - 3 раза уменьшить длину побегов в верхнем ярусе куста и повысить их морозоустойчивость (Zranger, Hague, 1968). Эффективное действие оказывает и хлорхолинхлорид.
Большие убытки в промышленном плодоводстве происходят вследствие подмерзания цветковых почек у косточковых культур. Практически у всех косточковых морозоустойчивость генеративных органов положительно коррелирует с более поздним началом их дифференциации и более медленными темпами их дальнейшего развития в осенний период. Замедления темпов развития цветковых почек и повышения их морозоустойчивости можно достичь с помощью регуляторов роста. Так, положительное действие оказывает применение гиббереллина! Эффективность этого препарата во многом зависит от сроков обработки. При поздних сроках опрыскивания деревьев, когда уже мало проявляется тормозящее действие гиббереллина на дифференциацию цветков, эффективность его снижается. Однако при ранних сроках применения он может оказывать отрицательное действие, вызывая слишком затяжной рост побегов, что приводит к снижению общей зимостойкости дерева и формированию малого количества цветковых почек (Edgerton, 1966). В последнее время положительные результаты получены при использовании этрела. Установлено достаточно эффективное его действие на повышение зимостойкости цветковых почек персика. Наибольшее влияние этрел оказывал при обработке растений в середине сентября, когда с деревьев был снят урожай (Proebsting, Mills, 1969).
Одним из приемов преодоления повреждающего действия весенних заморозков является задержка сроков цветения. Наибольшее значение эти приемы имеют при выращивании плодовых пород и сортов, цветение которых начинается в наиболее ранние сроки весной. Это относится к косточковым культурам, в первую очередь к абрикосу, а также к некоторым ягодникам. Задержка цветения имеет положительное значение и для других плодовых деревьев, в частности для яблони и груши. В связи с этим определенный интерес представляет использование с этой целью регуляторов роста.
Установлено, что гиббереллин задерживает не только начало дифференциации цветковых почек, но и темпы их развития, в том числе и микроспорогенеза (Corgak, Widmoyer, 1971). Так, обработка деревьев персика в первой половине августа (концентрация раствора 200 мг/л) на две недели задерживала наступление фазы цветения весной следующего года. При этом важно отметить, что под влиянием гиббереллина повышалась морозоустойчивость цветковых почек и до цветения. В ряде исследований показана положительная роль в задержке цветения и при обработке деревьев яблони и груши аларом и хлорхо-линхлоридом.
Регуляторы роста могут оказывать и непосредственное влияние на повышение морозоустойчивости цветков во время цветения. Установлено, что N-деценилянтарная кислота способствует повышению устойчивости цветков к отрицательной температуре. Так, при обработке этим препаратом деревьев яблони за 4 ч до начала похолодания удавалось повысить устойчивость цветков до - 6°С. Опрыскивание в другие сроки было неэффективным. Деценилянтарная кислота и некоторые ее моноамиды существенно повышают морозоустойчивость цветков земляники. Полагают, что положительное действие этого препарата происходит в результате проникновения его в липидные слои клеточных мембран, что способствует лучшей проницаемости их по отношению к воде (Kuiper, 1964).
Регуляторы роста (гиббереллин) применяют также с целью индуцирования завязывания и развития плодов, поврежденных заморозками. Исследования, проведенные в Нидерландах в 1965 - 1967 гг., показали, что многие сорта груши под влиянием гиббереллина способны формировать партенокарпные плоды после повреждения цветков заморозками. В результате обработанные деревья давали высокий урожай, а у контрольных он отсутствовал (Varga, 1969).
В последние годы с целью повышения морозоустойчивости плодовых культур начинают применять специальные вещества, получившие название криопротектанты. К ним относятся этиленгликоль, глицероль и др. Установлено, что применение криопротектантов в конце лета - начале осени позволяет повышать морозоустойчивость яблони до - 3°С.
Экологические факторы оказывают существенное влияние на повреждение цветков у плодовых растений весенними заморозками. Хорошо известна роль рельефа местности в усилении или ослаблении повреждающего действия заморозков. Это обусловливает необходимость тщательной оценки территории, отводимой под закладку сада, создания защитных лесонасаждений, водоемов. Важное значение имеют и специальные приемы борьбы с заморозками.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'