Особенности морфогенеза апекса побега у периодично плодоносящих деревьев яблони
Апекс побега, как уже отмечалось, в определенной степени является автономной саморегулирующейся биологической системой. Поскольку одной из главных физиологических функций этой части растительного организма является образование и детерминация зачатков будущих органов растения (листьев, побегов, цветков), то, естественно, изучение особенностей морфогенеза апекса занимает важное место в явлении периодичности плодоношения.
Рассматривая особенности морфогенеза апекса яблони, можно заметить существенные различия в этом процессе у деревьев с обильным урожаем (не способных к закладке цветков) по сравнению с деревьями без плодоношения (способных формировать цветковые почки) (Агафонов, Дмитриева, 1974).
У деревьев без урожая число зачатков листьев в апексе побегов, особенно кольчаток, всегда больше (табл. 4). При этом циклы листообразования протекают более напряженно во времени (начинаются на 2 - 3 недели раньше, чем у деревьев с урожаем). В данном случае ритм морфогенеза апекса у деревьев с обильным плодоношением идет по пути вегетативного развития. В то же время более напряженное протекание процессов, направленных на формирование метамерных органов, является, вероятно, одним из обязательных условий перехода апекса в генеративное развитие.
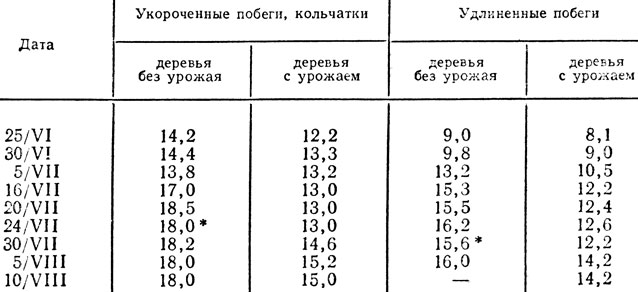
Таблица 4. Число зачатков листьев в апексе побегов яблони Осеннее полосатое
* (Начало образования видимых зачатков органов цветка.)
Количественные признаки апекса побега имеют непосредственную связь с переходом его к генеративному развитию. Проявляется это не только в числе пластохронов, но и в размере почки. У неплодоносящих деревьев размер почки и соответственно апекса всегда больше. При этом установлено, что в переходный период непосредственно перед началом образования органов цветка происходит значительное уменьшение диаметра основания конуса нарастания, что характерно вообще для высших растений (Серебрякова, 1963).
Следовательно, уменьшение диаметра основания конуса нарастания связано с началом анатомической перестройки апекса при переходе его к генеративному развитию и эта перестройка начинается после образования определенной критической массы меристематических клеток.
Анатомическое изучение апекса кольчаток показывает, что при переходе к репродуктивному развитию происходит цитогистологическая перестройка конуса нарастания. При этом заслуживает внимания факт, что эта перестройка, выражающаяся в уплощении конуса нарастания, исчезновении четких различий между клетками туники и конуса и увеличении числа мелких меристематических клеток, в определенной мере свойственна и апексу кольчаток деревьев с обильным плодоношением, хотя он и не переходит к генеративному развитию.
Приведенные данные позволяют полагать, что апекс кольчаток деревьев с обильным плодоношением также получает сигнал к включению механизма генеративного развития. Однако вследствие того, что размер апекса, прежде всего количество меристематических клеток и тканей, не достигает критической массы, дальнейшее развитие репродуктивного этапа задерживается. Следовательно, включение пускового механизма зависит, вероятно, от генотипа и онтогенетического состояния растения, в то время как полное завершение генеративного развития апекса, или реализация импульса цветения, в значительной мере определяется трофическими факторами.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'