Глава 5. Физиологические основы плодоношения
Возделывание культурных растений своей главной целью ставит получение максимально возможного урожая с единицы площади. В связи с этим особо важное значение приобретает изучение физиологических процессов вегетативного и генеративного развития, а также изучение вопросов, связанных с разработкой путей их оптимальной сбалансированности. Только на этой основе можно правильно разработать агротехнику культурных растений, позволяющую выявить их потенциальную продуктивность. Это одно из главных положений в интенсификации современного сельскохозяйственного производства.
В плодоводстве разработка путей оптимальной сбалансированности вегетативного и генеративного развития имеет особое значение. Решение этого вопроса необходимо для получения максимально возможного урожая, а также для создания скороплодных, ежегодно плодоносящих насаждений с высокой устойчивостью к неблагоприятным факторам среды в течение всего продуктивного периода их жизни.
Развитие высших растений представляет собой сложный физиологический процесс, критическим периодом которого является переход от вегетативного состояния к генеративному. Изучению механизма этого процесса в течение длительного времени уделяется значительное внимание в биологической и сельскохозяйственной науке.
Первым из исследователей, пытавшимся объяснить механизм перехода растений к генеративному развитию, был немецкий физиолог Ю. Сакс (1892). Он высказал гипотезу, что переход растений к цветению происходит под влиянием особых веществ. И хотя эта гипотеза не нашла экспериментального подтверждения, необходимо отметить одно правильное положение ее о том, что для перехода растений к генеративному развитию необходимы особые условия, отличные от тех, при которых происходит вегетативный рост. Гипотеза Сакса в дальнейшем получила новое освещение с последующей разработкой учения о ростовых веществах.
Значительный вклад в разработку учения о ростовых веществах внес советский физиолог М. X. Чайлахян (1937). Он высказал предположение, что для перехода растений к генеративному развитию необходимо накопление достаточного количества специфического гормона цветения, названного им флоригеном. Многочисленные опыты М. X. Чайлахяна и его сотрудников с прививками, а также обработка растений, не получивших соответствующей фотоиндукции, вытяжкой из растений, получивших это воздействие (Lincoln et al., 1962), казалось бы, убедительно подтверждали теорию М. X. Чайлахяна. Однако до настоящего времени еще не удалось обнаружить этот гормон в растениях.
Позднее М. X. Чайлахян (1958) высказал мнение о том, что предполагаемый гормон цветения является двухкомпонентиым (состоит из гиббереллина и гипотетического антезина). Первый определяет рост и развитие цветковых стеблей, а второй необходим для формирования органов цветка. Однако в настоящее время едва ли есть основание относить гиббереллин к специфическому гормону цветения. Его действие эффективно только для тех растений, у которых по тем или иным причинам (отсутствие фотоиндукции, яровизации) задерживается стеблевание. В то же время широко известно и отрицательное действие гиббереллина на генеративное развитие.
В последние годы доказана определенная роль цитокининов в генеративном развитии (Бутенко, 1964). Как известно, эти соединения существенно стимулируют органообразовательные процессы у растений (Кулаева, 1973).
Следовательно, приведенные данные, с одной стороны, не подтверждают специфичность предполагаемых гормонов цветения, а, с другой, указывают на то, что, по-видимому, существует более значительное число соединений, входящих в гормональный комплекс. Вероятно, контролирование цветения, как и вообще процесса развития в целом, определяется сбалансированностью различных регуляторов роста, входящих в гормональный комплекс. Причем немаловажное значение в этом комплексе должны иметь не только стимуляторы, но и ингибиторы роста, прежде всего природный ингибитор абсцизовая кислота (дормин). Эти вещества способствуют прекращению видимого роста побегов, что у многих древесных растений, в том числе и плодовых, является необходимым условием для перехода апекса* из вегетативного состояния в генеративное.
* (Термином "апекс" определяется не только собственно апикальная меристема, но и лежащая ниже субапикальная зона, характеризующаяся активной детерминацией зачатков листьев.)
В практике плодоводства давно было замечено, что многие приемы, тормозящие рост побегов (кольцевание и сгибание ветвей), способствуют закладке цветковых почек. Поскольку эти приемы способствуют накоплению в побегах углеводов, то, естественно, возникло предположение о важном значении этих веществ в переходе почки к генеративному развитию (Мюллер-Тургау, 1898).
Таким образом, углеводам отводилась роль своего рода цветообразующих веществ. Утверждалось также, что индуцирующее действие углеводов на процесс закладки цветков наступает при определенной концентрации Сахаров в клеточном соке (Loew, 1905).
Подобное объяснение механизма перехода апекса побега из вегетативного состояния в генеративное не могло объяснить сущность не только весьма сложного процесса развития растений вообще, но и роль углеводов в этом процессе.
Впоследствии гипотеза концентрации клеточного сока получила свое развитие в работах И. А. Коломийца (1966), проводившего свои опыты с плодовыми культурами. Оригинальными исследованиями И. А. Коломиец показал, что закладка цветковых почек у яблони зависит не от содержания углеводов (или углеводно-азотного соотношения), а от повышенной концентрации клеточного сока. Причем увеличения этой концентрации можно достичь разными путями: с помощью кольцевания побегов, что приводит к накоплению в клеточном соке углеводов, или путем внесения повышенных доз минеральных удобрений, способствующих накоплению азотистых соединений и влияющих на поглощение воды из почвенного раствора.
Предположение И. А. Коломийца представляет несомненный интерес в изучении процесса перехода растений от вегетативного развития к генеративному. Однако предстоит решить основной вопрос, каким путем изменение концентрации клеточного сока непосредственно затрагивает механизм развития апекса побега. Судя по опытам И. А. Коломийца, можно полагать, что повышение концентрации клеточного сока является необходимым условием для прекращения роста побега. Наряду с другими факторами (благоприятная температура, освещенность, достаточное снабжение элементами питания и пластическими веществами), это, вероятно, является важным физиологическим моментом, поскольку известно, что закладка органов цветка у многих плодовых и вообще древесных растений начинается именно только после прекращения роста побегов.
Прекращение роста побегов, очевидно, приводит к накоплению в их апексе наиболее важных функциональных соединений - нуклеиновых кислот и белков, а также способствует накоплению необходимого количества меристематических недетерминированных клеток в конусе нарастания. Таким путем апекс достигает определенной критической массы, что, видимо, является обязательным условием для перехода растения от вегетативного этапа развития к генеративному.
Нельзя исключить и того, что прекращение роста побегов оказывает определенное влияние на сбалансированность гормональных веществ. Возможно, при этом усиливаются функции гормонов, от которых в значительной мере зависит митотическая активность клеток конуса нарастания и ослабляются функции гормонов, способствующих росту клеток в фазе растяжения. Косвенно это подтверждается тем, что обработка плодовых деревьев синтетическими ингибиторами роста вскоре после прекращения роста побегов существенно стимулирует генеративное развитие (Тromр, 1973). Вероятно, в этом случае экзогенные регуляторы роста еще более усиливают изменение в сбалансированности гормональной системы в сторону индукции цветения. В пользу этих доводов говорят и опыты с изменением ориентации плодовых растений: помещение последних в горизонтальное положение после прекращения роста побегов также усиливает генеративные функции.
Торможение роста побегов у растений, находящихся в благоприятных условиях произрастания, как правило, стимулирует генеративное развитие. Это в первую очередь относится к плодовым породам, у которых цветковые почки закладываются большей частью на главной оси побега. Следовательно, процессы роста и развития, очевидно, нельзя отделять от сбалансированности гормональной системы, поскольку они взаимообусловлены (взаимосвязаны).
Значительный вклад в изучение механизма развития растений внес немецкий физиолог Клебс (1895). Он полагал, что процессы развития контролируются не только углеводами, но и в неменьшей мере неорганическими соединениями и прежде всего азотными. Выводы Клебса послужили основанием для предположения о том, что в переходе растений от вегетативного этапа к генеративному решающая роль принадлежит количественному соотношению между углеродом и азотом. При этом полагали, что для перехода к генеративному развитию необходимо высокое отношение углерода к азоту (Краус, Крейбилл, 1918).
Подобное упрощенное объяснение доктрины Клебса в сущности явилось дальнейшим развитием идей Мюллер-Тургау (1898) и, конечно, не могло быть достаточно удовлетворительным, хотя и надо признать, что в общем у растений закономерно повышается отношение углерода к азоту при переходе их к генеративному развитию. Однако при этом нужно иметь в виду, что в абсолютном выражении значительно увеличивается не только содержание углеводов, но и азотных веществ. Достаточно убедительно это было доказано работами американских исследователей (Поттер, Филипс, 1930). Не всегда высокое отношение углерода к азоту может способствовать переходу апекса вегетативного побега в генеративное развитие. Такое состояние может возникнуть при недостаточном обеспечении растений азотным питанием, но и в этом случае, несмотря на высокое отношение углерода к азоту, дефицит азотистых соединений может тормозить процесс формирования цветковых органов. Отрицательное действие недостатка азотистых веществ на генеративное развитие часто наблюдается у плодовых деревьев в годы с обильным плодоношением.
Главным недостатком гипотезы Е. Крауса и X. Крейбилла (1918) является то, что она не затрагивает сущности механизма развития растений. Очевидно, причины перехода растений от вегетативного этапа к генеративному надо искать не в простом отношении углерода к азоту, а в процессах метаболизма углеродов и азотистых соединений и в процессе обмена веществ вообще, выделяя при этом центральные вопросы.
Понимание механизма генеративного развития только с позиций отношения углерода к азоту привело к упрощенному пониманию гипотезы Г. Клебса. Поскольку закономерности в отношениях между углеродом и азотом не всегда соответствуют переходу растений от вегетативного типа к генеративному, то это нередко приводит к тому, что представление Клебса считается ошибочным (Туркова, 1967).
Однако рациональное зерно в учении Клебса состоит не в простом отношении углерода к азоту, которое можно изменить условиями питания, а в том, что проявление генетического потенциала растений находится в определенной зависимости от факторов внешней среды. При этом Клебс предполагал, что влияние внешней среды на переход растений к генеративному развитию осуществляется путем воздействия на процессы, протекающие непосредственно в клетках. Он считал также, что одним из основных показателей проявления генетического потенциала может быть определенный состав клеточных белков.
Таким образом, хотя Клебс и предполагал, что в механизме развития растений большая роль принадлежит отношению углерода к азоту, однако решающее значение он придавал все-таки взаимодействию генотипа и внешней среды. Только при таком понимании учение Клебса Могло получить правильное развитие как в теоретическом, так и в практическом плане. В частности, разрабатывая это учение, нельзя не принимать во внимание такие факторы, как возрастное состояние растительного организма, реакция растений на фотопериодические и температурные воздействия, а также не учитывать физиологические функции основных органов растений - корней и листьев.
Только с учетом всего комплекса внутренних и внешних факторов гипотеза Клебса может объяснить отдельные приемы, применяемые в сельскохозяйственной практике. Так, в зависимости от фенофаз развития плодового дерева азотные удобрения могут оказывать противоположное действие: применение их в период роста побегов задерживает закладку цветковых почек, а после окончания роста побегов усиливает этот процесс.
В настоящее время известно, что белки составляют метаболическую основу клетки, определяя ее форму и функции. Состав этих белков контролируется геномом, дифференциальная активность которого, вероятно, зависит от гормональных веществ. Следовательно, оказывая влияние на эндогенную регуляторную систему условиями внешней среды, мы можем изменять и метаболическую направленность, и активность клетки и таким путем оказывать непосредственное воздействие на развитие всего растения. Можно полагать, что сказанное лежит в основе современной разработки гипотезы Клебса и является теоретической предпосылкой для применения физиологически активных веществ в плодоводстве.
Поскольку бесспорна решающая роль генома в развитии растительного организма, то, естественно, в последние годы большое внимание уделяется изучению метаболизма нуклеиновых кислот в связи с генеративным развитием. Высказывается предположение, что для перехода конуса нарастания побега из вегетативного состояния в генеративное необходимо прежде всего накопление в верхушечной меристеме ДНК (Агафонов и др., 1970). Основано это на том, что началу формирования органов цветка в конусе нарастания предшествует накопление массы недетерминированных меристематических клеток, а сам процесс начинается повышением митотической активности этих клеток (Серебрякова, 1963). И хотя синтез ДНК еще нельзя считать пусковым механизмом митоза, однако ее накопление является необходимым условием для прохождения митотических циклов (Цанев, Марков, 1964).
Роль нуклеиновых кислот в процессе перехода растений к цветению показана многими исследователями. Установлено, что чем более благоприятные условия для генеративного развития, тем раньше достигается максимум их содержания в верхушечных меристемах (Туркова, 1967). В то же время в условиях, тормозящих генеративное развитие (недостаток света, избыток азота и другие факторы), наблюдается снижение содержания нуклеиновых кислот. При этом важно отметить, что содержание ДНК понижается более значительно, чем РНК.
Убедительно показана роль нуклеиновых кислот на примере использования антиметаболитов нуклеинового обмена. Было установлено, что эти вещества вызывают торможение зацветания растений, получивших необходимую фотопериодическую индукцию (Bonner, Leebart, 1962).
Высказывается предположение, что переход растений к генеративному развитию зависит от специфичной РНК, отличающейся по нуклеотидному составу: несколько другое отношение между гуанином и аденином, чем в РНК, участвующей в морфогенезе вегетативных органов (Haas, 1961). Следовательно, это предположение позволяет считать, что процесс перехода апекса побега от вегетативного этапа развития к генеративному связан с системой: специфичные гены ДНК - специфичная РНК - специфичный белок.
Отмечая большую роль нуклеиновых кислот в процессе зацветания растений, необходимо иметь в виду, что накопление этих соединений требует больших затрат энергии. Отсюда очевидно, что переход растении к генеративному развитию может осуществляться только при высоком энергетическом уровне. Наоборот, в условиях, неблагоприятствующих процессам фосфорилирования, наблюдается снижение накопления нуклеиновых кислот и этим тормозится процесс зацветания (Туркова, 1967).
Успешное изучение механизма развития растений невозможно без применения количественного анатомо-морфологического анализа. В этой связи особого внимания заслуживает изучение морфогенеза апекса побега, являющегося в определенной степени саморегулирующейся биологической системой и занимающего ключевое положение в процессах вегетативного и генеративного развития растений. В настоящее время многочисленные работы позволяют довольно четко представить анатомо-морфологическую структуру апекса побега у покрытосемянных растений (Серебрякова, 1963, и др.). Строение апекса побега у плодовых также соответствует этой структуре (рис. 21).
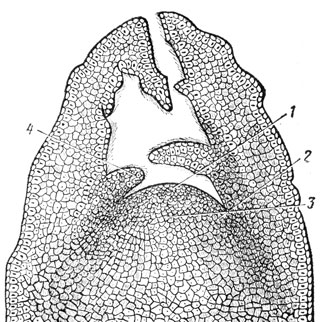
Рис. 21. Апекс побега яблони Осеннее полосатое: 1 - туника; 2 - инициальное кольцо (периферическая зона); 3 - корпус; 4 - зачаток листа
Функциональная деятельность клеток и тканей разных зон апекса неодинакова. В период вегетативного развития побега наибольшей митотической активностью обладает периферическая зона (клетки инициального кольца), где происходит процесс листообразования.
С наступлением префлорального периода активность периферической меристемы снижается, что приводит к прекращению процесса листообразования. Одновременно происходит нарастание массы собственно конуса нарастания: увеличивается количество слоев туники и количество меристематических недетерминированных клеток этой зоны. При этом сглаживаются различия между клетками туники и корпуса; клетки последнего перестраиваются, приобретая свойства периферической меристемы, то есть становятся мелкоклетными, и повышается их митотическая активность. Таким путем клетки центральной зоны корпуса вместе с туникой образуют мелкоклетную мантию вокруг разрастающегося паренхимного ядра. Морфологически переход апекса в генеративное развитие характеризуется значительным уменьшением диаметра основания собственно конуса нарастания, прилегающего к инициальному кольцу, но при этом увеличивается длина конуса.
Отмеченная анатомическая перестройка свойственна префлоральному периоду в развитии апекса и заканчивается начальным этапом в образовании непосредственно органов цветка: удлинение конуса нарастания представляет собой формирование оси соцветия. Таким образом, в процессе перехода апекса побега от вегетативного этапа развития к генеративному происходит существенное перераспределение цитогистологических зон, изменяется митотическая активность клеток верхушечной меристемы и их органообразовательная деятельность.
Переход апекса к генеративному развитию имеет непосредственную связь с количественными признаками побега. Так, формирование органов цветка начинается только после образования определенного количества зачатков листьев или после прохождения определенного числа пластохронов* (Сабинин, 1963; Серебрякова, 1963).
* (Пластохрон - биологическая единица времени, характеризующаяся образованием одного узла в апексе или протекающая в период между началом образования одного зачатка листа до начала образования другого зачатка.)
Количество листьев на побеге является устойчивым наследственным признаком у травянистых растений. Однако у многолетних древесных видов, в том числе и у плодовых, имеющих сложную структуру надземной системы, этот признак зависит не только от вида, но и от разнотипности ветвей и побегов. В данном случае более устойчивым признаком, вероятно, может быть не количество листьев на сформировавшемся побеге, а число зачатков листьев в апексе, поскольку есть основания полагать, что процесс образования зачатков листьев у древесных пород протекает более стабильно и в меньшей мере зависит от факторов внешней среды.
В плодоводстве известно, что формирование цветковой почки начинается только после образования определенного числа зачатков листьев (Ро, 1929). В условиях Московской области переход апекса кольчатки в генеративное развитие у яблони Осеннее полосатое начинается после прохождения 18 пластохронов, а у Антоновки - после 15 пластохронов (Агафонов, Дмитриева, 1973). Однако число пластохронов, определяющее переход апекса в генеративное развитие, зависит не только от сорта или вида, но и от типа побега. Так, в отличие от кольчаток образование органов цветка на удлиненных побегах яблони Осеннее полосатое начинается после 15 пластохронов.
Следовательно, при оценке количественной зависимости как эволюционного признака между образованием зачатков метамерных органов и переходом побега к генеративному развитию у древесных пород необходимо принимать во внимание и разнотипность побегов. Тем не менее отмеченные особенности в развитии апекса убедительно подтверждают известное положение Д. А. Сабинина (1963) о том, что узел с междоузлием, в том числе применительно к апексу побега, является элементарным этапом в развитии растений.
Процесс формирования зачатков листьев в апексе побега яблони протекает достаточно стабильно в разных условиях произрастания (Агафонов, Дмитриева, 1973). Однако было бы неправильным считать, что прохождение необходимого числа пластохронов уже достаточно для перехода побега к генеративному развитию. В условиях крайне жаркого и сухого лета 1972 г. в апексе удлиненных побегов яблони Осеннее полосатое сформировалось необходимое количество зачатков листьев. Между тем в отличие от прошлого года (достаточно влажное и нежаркое лето), когда после такого же числа пластохронов (15) начался процесс образования органов цветка, в 1972 г. формирование цветковых почек на удлиненных побегах не наступило.
Приведенные данные указывают на то, что наряду с наследственными свойствами в процессе морфогенеза апекса побега определенное значение имеют факторы окружающей среды. При этом отмеченные особенности в характере развития апекса побега дают основание полагать, что генеративный этап развития в большей мере зависит от условий произрастания, чем вегетативный.
Последний довод находит подтверждение и в особенностях развития растений под влиянием фотопериодических и температурных индукций (растения короткодневные и длиннодневные, яровые и озимые). В этом случае переход растений к генеративному развитию происходит только под воздействием соответствующей индукции или в определенных условиях произрастания.
В настоящее время еще не выяснен весь механизм действия указанных индукций на развитие растений. Можно полагать, что они способствуют прохождению физиолого-биохимических и анатомо-морфологических изменений, проявляющихся в накоплении определенной критической массы, необходимой для перехода организма в новое качественное состояние. Однако каким путем проявляется непосредственно сигнал (импульс) этой индукции, включающий пусковой механизм генеративного развития, еще неизвестно.
Вполне возможно, что один из путей стимулирования цветения связан с деятельностью ферментов, оказывающих определенное влияние на сбалансированность гормональной системы. Наиболее вероятен указанный путь для температурной индукции. Что касается влияния фотопериода, то здесь, видимо, большая роль должна принадлежать пигментной системе растительного организма и прежде всего фитохрому (Туркова, 1967).
В переходе растительного организма к генеративному развитию решающее значение имеет его возрастное состояние. Образование органов цветка у растений может начаться только после прохождения ювенильного этапа онтогенеза. Продолжительность этого этапа у растений различна. У травянистых, особенно однолетних, она может быть очень короткой и исчисляться днями, у многолетних древесных может продолжаться в течение нескольких лет. В плодоводстве отмеченное обстоятельство важно учитывать при разработке различных приемов, индуцирующих формирование цветковых почек. Так, применение физиологически активных веществ, ускоряющих начало плодоношения, не может быть эффективным при обработке сеянцев яблони в ювенильный период их развития (Jonkers, 1973). В этой связи представляет интерес схема развития плодовых деревьев и вообще растений, предложенная Циммерманом (Zimmerman, 1973). Как полагает автор, стимулирование цветения становится возможным только при наступлении промежуточной фазы развития.
Рассмотренные анатомо-морфологические и физиолого-биохимические особенности у растений в связи с их переходом в цветковоспелое состояние указывают на то, что в изучении этого процесса достигнуто многое. Однако следует признать, что сущность этого явления до настоящего времени остается полностью не раскрытой.
Современные представления о переходе растений к генеративному развитию позволяют выразить этот процесс следующим образом.
Ювенильный период: усиленный вегетативный рост, преобладание функций корня и гиббереллинов в гормональной системе.
Промежуточный период: постепенное накопление критической массы нуклеиновых кислот, белков, меристематических клеток в конусе нарастания, постепенное затухание ростовых процессов, сбалансирование функций корня, листа и гормональной системы.
Цветковоспелый период: преобладание функций листа и ингибиторов в гормональной системе, критическая масса достигла необходимой величины, включение пускового механизма по системе: начало активности специфичных генов ДНК - синтез специфичной РНК - синтез специфичных белков - анатомо-морфологическая органообразовательная перестройка апекса побега с переходом во флоральное развитие.
Можно полагать, что у многолетних древесных растений включение пускового механизма цветения (импульс цветения) возможно только при наступлении промежуточного периода в развитии организма. На этом этапе становится достаточно эффективной разработка различных приемов, ускоряющих вступление в продуктивный период плодовых деревьев.
Достижения науки последних лет в области физиологии развития растений уже теперь легли в основу разработки многих агротехнических приемов возделывания сельскохозяйственных растений. Особенно наглядно это видно на примере с плодовыми культурами. В настоящее время с помощью физиологически активных веществ (регуляторы роста) с большим успехом разрабатываются приемы направленного воздействия на процессы вегетативного и генеративного развития плодовых и ягодных растений. Появляется реальная возможность контролировать рост и плодоношение, создавать насаждения с максимально возможным потенциалом продуктивности, регулярным плодоношением и устойчивые к неблагоприятным факторам среды.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'