Факторы, определяющие оптимально-продуктивный размер и форму кроны
Разнокачественность вегетативных и репродуктивных органов в корне дерева. Характерной особенностью плодоносных органов у плодовых деревьев является их физиологическая и биологическая разнокачественность, причиной которой могут быть условия питания, освещенности, водного режима и некоторые другие факторы, неодинаковые в разных участках кроны. Разнокачественность плодоносных ветвей определяет количество и качество урожая, формирующегося на них. Старые и менее развитые плодоносные органы утрачивают свою жизнеспособность и становятся менее продуктивными. Следовательно, при разработке системы формирования и обрезки необходимо добиваться, чтобы крона плодового дерева была наполнена хорошо развитыми плодоносными ветвями и побегами.
Важным биологическим свойством плодоносных органов, которое крайне необходимо учитывать при разработке системы формирования и обрезки, является их долговечность. Большая часть плодоносных образований в кроне плодового дерева отмирает после 8-летнего возраста. А поскольку жизнь плодового дерева более продолжительна, то это приводит к оголению основных ветвей вследствие центробежного перемещения плодоносных образований. В результате у взрослого дерева периферийные и центральные участки кроны становятся неравноценными по своей продуктивной способности. Следовательно, при определении наиболее оптимальных размеров кроны важно учитывать соотношение между оголенной частью кроны и заполненной продуктивными ветвями.
При определении оптимального размера кроны важно также учитывать и тот факт, что плодовые деревья на соответствующих этапах онтогенеза обладают способностью омолаживаться. Наряду с процессами старения и отмирания ветвей в кроне возникает процесс и их возобновления, который чаще всего проявляется в образовании молодых ветвей у основания скелета кроны.
Как правило, процесс омоложения начинается в тот период развития плодового дерева, когда у него наблюдается преобладание усыхания обрастающих ветвей над процессом новообразования, то есть формирования новых обрастающих ветвей на периферии кроны. С этого периода начинается затухание жизненного потенциала плодового дерева, что проявляется в прогрессивном уменьшении ростовых процессов.
Образование молодых ветвей у основания главных в конце концов приводит к тому, что начинается более интенсивное отмирание не только обрастающих ветвей на периферии кроны, но и выпадение более крупных частей кроны - основных ветвей первого и второго порядка. В связи с этим возникшие у основания кроны молодые ветви начинают формироваться в главные, на которых появляются обрастающие плодоносные органы, в результате возникает новый цикл развития кроны дерева.
Указанные закономерности в развитии основных и обрастающих ветвей в кроне плодового дерева послужили основанием П. Г. Шитту для разработки закона циклической смены скелетных и обрастающих частей у плодовых деревьев и кустарников (см. главу 3).
Разработанный П. Г. Шиттом (1952) закон циклической смены скелетных и обрастающих частей имеет исключительно большое значение в теории и практике формирования и обрезки плодовых деревьев. Без знания этого закона нельзя разработать оптимально- продуктивные размеры крон и их форму, нельзя правильно определить оптимальную продолжи-тельность продуктивного периода жизни плодового дерева. При этом следует отметить, что правильное применение закона циклической смены обрастающих и скелетных частей на практике возможно только с учетом двух главнейших факторов. Во-первых, темпы и характер перемещения вегетативных и плодоносных органов в кроне плодового дерева определяются наследственными свойствами породы или сорта, во-вторых, эти свойства довольно успешно поддаются регулированию агротехническими приемами, что позволяет изменять не только количество циклов, но и их динамичность, то есть продолжительность во времени. Нельзя не отметить и третий фактор, имеющий большое практическое значение. Определяя на основе закона циклической смены ветвей оптимально-продуктивный размер кроны и продолжительность продуктивного периода жизни плодового дерева, необходимо руководствоваться прежде всего потенциалом продуктивности* плодовых насаждений.
* (Объем продуктивной части кроны в расчете на единицу площади сада.)
Световой режим кроны дерева и оптические свойства листьев. Центробежное перемещение продуктивных органов в кроне дерева, как уже отмечалось, является следствием процесса их развития (старения). Однако темпы развития зависят от многих факторов. На этот процесс, кроме наследственных причин, большое влияние оказывают условия окружающей среды. Так, продолжительность жизни продуктивных органов нередко зависит от подмерзания в неблагоприятные зимы.
Процесс старения и отмирания ветвей у плодовых деревьев в значительной мере зависит и от светового режима в кроне дерева. Плодовые породы в основном относятся к светолюбивым растениям. Следовательно, с уменьшением освещенности будет наблюдаться расстройство фотосинтезирующей системы дерева, что приведет к преждевременному старению и отмиранию обрастающих ветвей.
С увеличением возраста деревьев и их размера существенно снижается освещенность внутренних участков кроны. В настоящее время установлено, что количество ФАР заметно снижается с продвижением к центру кроны. Так, на расстоянии 1,0 - 1,5 м от периферии проникает 30 - 50% ФАР, а значительная часть кроны получает менее 30% ФАР (рис. 39). Согласно данным Кэйна (Cain, 1971), нормальное функционирование плодоносных органов, прежде всего цветковых почек, возможно только при поступлении ФАР в количестве не менее 50% дневной.
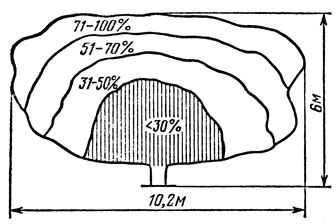
Рис. 39. Проникновение физиологически активной радиации в разные участки кроны крупногабаритного дерева яблони Мекинтош. По Луней
Указанные особенности освещенности разных участков кроны у плодовых деревьев дают основание полагать, что слабое проникновение ФАР в центр дерева является одной из основных причин оголения главных ветвей. Вероятно, уменьшение солнечной радиации приводит к снижению функциональной деятельности листьев и в целом обрастающей ветви, что неизбежно должно способствовать ускорению процесса их старения и отмирания.
Уменьшение физиологической роли листьев, размещенных на более взрослых участках основных ветвей (ближе к центру кроны), подтверждается исследованиями оптических свойств листьев. Установлено, что наибольшей способностью к поглощению ФАР обладают листья, сформированные на более молодом возрастном участке скелетной ветви, то есть ближе к периферии кроны (Колесников, Агафонов, Хрыпова, 1971). В то же время не установлены существенные различия в оптических свойствах листьев, сформированных на разных типах плодоносных побегов. Следовательно, оптические свойства листьев определяются главным образом условиями освещенности.
На оптические свойства листьев большое влияние оказывают особенности их анатомического строения. Так, исследованиями В. А. Колесникова, Н. П. Соколовой и С. П. Гхош (1973) было установлено, что с увеличением возраста плодоносных органов (кольчаток и плодушек) быстро уменьшается толщина столбчатого мезофилла листьев. Кроме этого, листья более молодых обрастающих ветвей содержат больше хлорофилла в расчете на единицу площади. Отмеченные особенности листьев, очевидно, и определяют различия в их оптических свойствах.
Различия в оптических свойствах листьев оказывают существенное влияние на фотосинтетическую продуктивность. Установлено, что в листьях, размещенных на периферийной части кроны, интенсивность фотосинтеза в 2,5 раза выше по сравнению с размещенными в глубине кроны. При этом важно отметить, что различия в фотосинтезе листьев, размещенных на расстоянии 1,5 м от периферии и в центре (3 м от периферии) кроны, незначительны (Кудрявец и др., 1970; 1972). Следовательно, интенсивность фотосинтеза резко падает с продвижением от периферии кроны к центру.
Изменения в фотосинтетической продуктивности листьев зависят и от других причин. В частности, уменьшение освещенности затрагивает механизм устьичного аппарата. Так, у древесных растений в нижней части кроны с уменьшением освещенности увеличивается сопротивление устьиц (Turner, 1969). При этом показано, что свет является важным фактором в механизме сопротивления устьиц. Возраст листа оказывал меньшее влияние на этот процесс. Исходя из этого, можно полагать, что одной из причин уменьшения интенсивности фотосинтеза является увеличение устьичного сопротивления, препятствующее поглощению углекислоты.
Различия в метаболической активности листьев, размещенных в разных участках кроны плодового дерева, достаточно наглядно видны по содержанию Сахаров и свободных аминокислот в плодоносных побегах. На протяжении всего периода вегетации количество Сахаров в побегах, расположенных на периферии кроны, намного больше, чем в побегах, размещенных внутри кроны (Агафонов, Хрыпова, 1972). Очевидно, это связано с более эффективной работой ассимиляционного аппарата, размещенного на периферии кроны. В результате происходит более интенсивный синтез Сахаров, в первую очередь транспортных и наиболее реактивных форм (сахарозы, глюкозы), перемещающихся затем в другие органы плодового дерева.
Существенны различия и по содержанию аминокислот. Так, в побегах на периферии кроны в течение всего вегетационного периода их намного больше, чем внутри кроны.
Отмеченные особенности в содержании свободных аминокислот и Сахаров в разных частях кроны плодового дерева связаны с оптическими свойствами этих зон кроны, а также с различной фотосинтетической деятельностью листьев. Очевидно, вследствие этого в глубине кроны по сравнению с периферией существенно снижается синтез не только Сахаров, но и аминокислот, поскольку свет играет значительную роль в образовании последних (Кретович, Каган, 1967). Стимулирующее влияние света, вероятно, является одной из причин более интенсивного синтеза на свету безазотистых предшественников аминокислот и образования АТФ, необходимой как источник энергии для амидирования дикарбоновых аминокислот до соответствующих амидов.
Таким образом, более высокое содержание в плодоносных побегах на периферии кроны таких высокореактивных соединений, как сахара и аминокислоты, дает основание говорить о том, что в этих участках кроны имеются более благоприятные условия для метаболизма и, следовательно, для процессов роста и развития.
Особенности размещения продуктивных органов в кроне. Изучение особенностей формирования и размещения продуктивных органов (листьев и плодов) в кроне плодового дерева имеет первостепенное значение при разработке системы формирования и обрезки. Только на этой основе можно правильно рассчитать оптимальные параметры плодового дерева, позволяющие получать максимально возможный урожай с единицы площади насаждения.
Выше отмечалось, что с возрастом плодового дерева в его кроне происходит центробежное перемещение обрастающих ветвей. Так, у яблони оптимальный возраст плодоносных ветвей обычно не превышает 8 лет, после чего начинается процесс их интенсивного отмирания. Следовательно, у взрослых плодовых деревьев многолетние основные ветви, составляющие скелет кроны дерева, будут неравноценны с точки зрения размещения плодов и листьев на разных возрастных участках.
Исследования В. А. Колесникова, Н. В. Агафонова и Н. X. Хрыповой (1970) показывают, что в условиях Московской области у 15- 30-летних деревьев яблони на периферийной части главных ветвей, возраст которых менее 8 лет, размещается около 90% листьев. Длина этой части главных ветвей не превышает, как правило, 1,7 - 1,8 м, что составляет около 40% их общей длины у 30-летних и около 50% у 15-летних деревьев. Как видим, даже у сравнительно молодых деревьев яблони, находящихся в оптимальном возрастном периоде, значительная часть главных ветвей не несет на себе продуктивных органов.
Отмеченные особенности в размещении листьев по длине главных ветвей свойственны и генеративным органам. Основная часть цветков (около 80%) у взрослых деревьев яблони также размещается на периферийной части основных ветвей не старше восьми лет. Вполне естественно, что это приводит и к неравномерному размещению урожая по длине этих ветвей. Так, у 30-летних деревьев яблони на участке ветви моложе восьми лет располагается около 80% всего урожая дерева (табл. 35). Плоды, сформированные в этой зоне, отличаются более крупными размерами и лучше окрашены по сравнению с теми, которые размещены в более глубоких участках кроны.
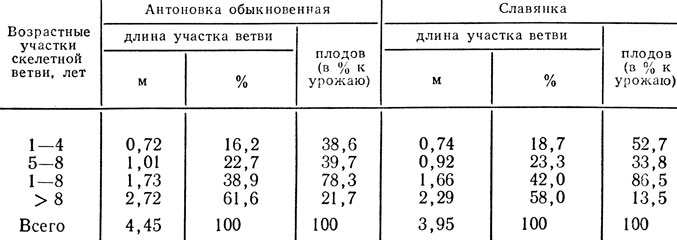
Таблица 35. Размещение урожая в кроне по возрастным участкам главных ветвей 30-летних деревьев яблони
Таким образом, у взрослых плодовых деревьев значительная часть главной ветви, а следовательно, и значительный участок кроны представляют собой оголенную зону и практически не участвуют в формировании урожая. Исходя из этого становится очевидной необходимость разделения кроны на две зоны - продуктивную и непродуктивную. Это следует учитывать при разработке системы формирования кроны, поскольку соотношение между этими зонами будет оказывать значительное влияние на потенциал продуктивности насаждения. Это тем более необходимо иметь в виду, если принять во внимание, что соотношение между продуктивной и непродуктивной зонами в кроне существенно изменяется во времени и может направленно регулироваться с помощью различных приемов, и прежде всего с помощью обрезки и формирования.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'