4.2. Роль окислительных ферментов и их изоферментов в проявлении нематодоустойчивости
Нематодоустойчивость растений может обусловливаться морфологическими особенностями устойчивых растений, образованием у них защитных некрозов, а также отсутствием возможности гидролизовать .ткани хозяина ферментами фитонематод, накоплением токсичных продуктов в пораженных, тканях и другими факторами (Мюге, 1965; Giebel, 1974). Существует мнение, что в ответ на внедрение нематод устойчивое растение высвобождает вещества, убивающие паразита. В другом случае устойчивость обусловлена тем, что растение-хозяин не продуцирует веществ, необходимых для воспроизведения нематод, и поэтому не привлекает их (Giebel, 1979).
В концепции о подразделении иммунитета к галловым нематодам на активный и пассивный представляет интерес активный, при котором личинки галловых нематод проникают как в восприимчивые, так и устойчивые растения. Однако в последних развитие личинок угнетается либо приостанавливается совсем, т.е. в результате активного иммунитета растение не в состоянии быть хозяином" на котором может продолжаться воспроизводство потомства нематод (Устинов,1959; Дементьева, Садакин, 1982).
Б.А. Рубин и Е.В. Арциховская (1965), развивая учение академика Х.Н. Баха о возможном участии оксидаз в защитных реакциях растительного организма, считают важным фактором естественного неспецифического иммунитета окислительные процессы, протекающие в тканях растений. Наиболее устойчивы к влиянию токсинов паразита ферменты растений пероксидаза и полифенолоксидаза, деятельность которых, в свете современных данных, связана друг с другом функционально. У иммунных форм растений инфекция в подавляющем большинстве случаев приводит к активированию этих ферментов. Последнее обусловлено новообразованием дополнительных количеств белков, обладающих каталитической активностью (Окопный, 1976).
Ферментативная система растения-хозяина, в частности система ферментов, участвующая в процессах дыхания, способна активно реагировать на внедрение паразита усилением метаболизма в зоне поражения. Эта реакция не является локальной, а распространяется на значительное расстояние от места поражения, причем активизируется не только деятельность оксидаз, но и вся сложная система реакций, участвующих в метаболических процессах (Бодрова, 1961).
Многие авторы, как указывалось выше, отводят важную роль окислительно-восстановительным ферментам. Основная роль оксидаз определяется их участием в дыхательных процессах, разрушении перекисных соединений, окислении моно- и полифенолов. Ослабление пероксидазной активности окислительных процессов и приводит к повышенного содержанию индолилуксусной кислоты. Последняя в случае поражения растения галловыми нематодами вызывает приток пластического материала в зону поражения и способствует разрастанию больных тканей, т.е. образованию галлов (Слепян, 1973).
Литературные данные об участии каталазы в патологическом процессе, вызванном инфекционным патогенном, весьма противоречивы. Некоторые авторы указывают на активное участие каталазы в реакциях сопротивляемости растений к инфекционным болезням (Пересыпкин, Шелехова, 1965; Кондратюк, Гоцуляк, 1969). Что касается пероксидазы, полифенолоксидазы и некоторых других ферментов, то в пораженной ткани растений их активность повышается (Мюге, 1965; Окопный, 1976, 1979; Maine" Keiman, 1960). Увеличение активности происходит в устойчивых и восприимчивых растениях, причем в последних активность возрастает больше, чем в устойчивых (Подкина, Погорелов, 1975).
Окислительная система растения связана не с дыханием, а с защитной функцией и естественным иммунитетом. При повреждении растения под действием полифенолокоидаз и пероксидаз образуются хиноны, создающие среду, препятствующую дальнейшему развитию внедрившегося паразита (Прокошев, Баранова, 1959; Вердеревский, 1968).
Отличительной особенностью устойчивых форм растений является присущая им высокая стабильность элементов окислительных систем, регулирующих процессы фото- и окислительного фосфорилирования. Исследование активности полифенолоксидазы корней устойчивых и восприимчивых к нематоде образцов позволило установить, что превращение фенольных соединений в корнях восприимчивых форм при поражении их мелойдогинами связано с повышением активности полифенолоксидазы. Это происходит либо за счет освобождения данного фермента при поражении клеток растительной ткани, либо освобождением метаболитов из клеток инвазированных нематодами, косвенно влияющих на повышение ее активности. Таким образом, полифенолоксидаза в корнях томата совместно с фенольными соединениями играет важную роль в проявлении реакций сверхчувствительности.
В корнях сортов и форм томата, контрастных по устойчивости к галловой нематоде, нами обнаружены количественные и качественные отличия изоферментов полифеноксидазы (рис. 8). Установлено, что изоферменты полифенолоксидазы митохондрий из корней томата устойчивых форм сохраняют в онтогенезе устойчивые компоненты со значением Rf 0,32 - 0,35, тогда как изоферменты полифенолоксидазы митохондрий из корней восприимчивых форм не содержат компонентов с подобным значением Rf. Также отсутствуют у восприимчивых образцов белковые компоненты с Rf, , близким к 0,84 - 0,85, тогда как в устойчивых они сохраняются в онтогенезе. Следовательно, устойчивые и восприимчивые к мелойдогинозу линии томата различаются между собой по числу изоферментных спектров и электрофоретической подвижности, что видно из их расположения на электрофореграммах.
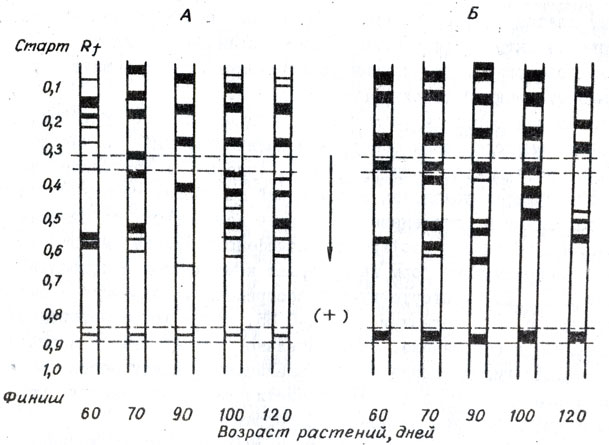
Рис. 8. Электрофореграммы изоферментов полифенолоксидазы митохондрии корней восприимчивого (А) и устойчивого (Б) к M.incognita сортов томата в онтогенезе
Полученные результаты позволяют высказать предположение, что полифенолоксидаза и ее изоферменты играют важную роль в механизме защитных реакций устойчивых форм растений, участвуя в окислении фенолов и образуя некрозы в местах поражения. Различия по активности полифенолоксидазы и числу ее изоферментов могут быть использованы как исходные критерии для оценки томата на устойчивость к галловой нематоде.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'