Ботаническая характеристика и особенности развития ячменя
Корни ячменя мочковатые - зародышевые (первичные) и узловые (вторичные). Зародышевыми называются потому, что в зачаточном виде они имеются в зародыше семени. При прорастании главный корень прорывает оболочку семени. Из зачатков на стебельке зародыша по очереди появляются еще 2 - 6 придаточных и в период между развитием второго и третьего листа - 2 колеоптильных (иногда 3 - 4) из колеоптильного узла.
В нашем опыте при проращивании зерновок ячменя сорта Донецкий 650 (каждая весом 52 - 54 мг) на глубине 7 см при температуре 16 - 19 °С и влажности 20 - 26% установлена следующая очередность появления зародышевых корней: через двое суток после посева образовались главный и 2-4 боковых, а через четверо суток - 5 - 6 зародышевых корней. Длина главного корня достигала через двое суток после посева 7 мм, через четверо суток 88 мм и через десять суток 118 мм, а боковые корни соответственно: первая пара - 5; 53 и 73 мм; вторая пара - 1,5; 23 и 56 мм; третья пара через четверо суток - 8 и через десять суток - 33 мм. На десятые сутки длина первого зеленого листа достигала 238 мм. На зародыше ячменя после 6-часового намачивания можно найти 5 - 8 обозначившихся корешков и, кроме того, несколько неясно видных корешков, которые развиваются лишь тогда, когда первые погибают.
Количество развитых зародышевых корней ячменя зависит и от уровня почвенного плодородия. Нами выявлено, что при улучшении условий минерального питания путем внесения удобрения у растений ячменя в фазе двух листьев было 6 зародышевых корней, а без удобрений - 4,5.
Количество зародышевых корней, образующееся во время прорастания ячменя, обусловлено качеством семян, в частности крупностью, сортовыми особенностями и условиями их формирования и созревания. В наших опытах на Эрастовской опытной станции семена урожая 1964 г., сформировавшиеся в верхнем ярусе колоса (в 18-м колоске, считая снизу), образовали несколько меньшее число зародышевых корней; они отличались меньшим весом, чем семена нижнего (3-го колоска) и среднего (10-го колоска) ярусов колоса (табл. 1).
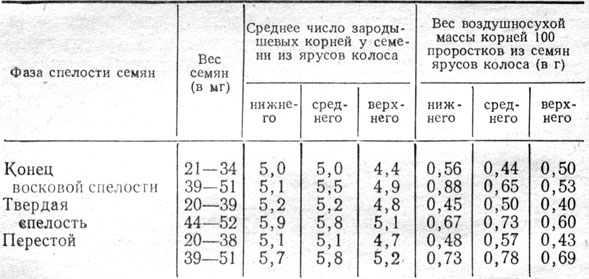
Таблица 1. Число зародышевых корней и их вес на 10-й день проращивания семян ячменя сорта Южный в зависимости от их веса, фазы спелости и места в колосе
Чем мельче семена, тем меньше корней и ниже их вес. Число корней у многорядного ячменя скорее определяется тяжеловесностью семян центральных или боковых колосков, чем местом формирования их по ярусам в колосе. Подтверждается это данными проращивания семян урожая 1963 - 1964 гг. (табл. 2).
При одинаковом весе семян центральных колосков любого яруса колосьев главных побегов, побегов кущения и нераскустившихся растений образовалось 5,2 - 6,8 зародышевого корня и вес их у 100 проростков составил 0,95 - 1,06 г. Меньше зародышевых корней с меньшим их весом образовалось у семян, сформировавшихся в боковых колосках, которые отличались значительно меньшим весом, чем в центральных колосках.
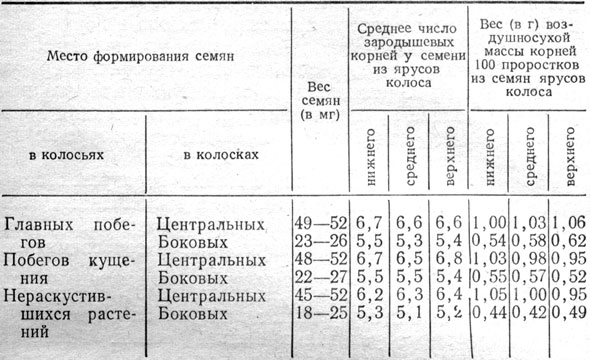
Таблица 2. Число зародышевых корней и их вес на 10-й день проращивания семян ячменя Грушевский в зависимости от веса и места в колосе
На количество зародышевых корней определенным образом сказывается и спелость зерновок. При высеве свежеубранных семян у зерновок, убранных в фазе твердой спелости, образовалось больше зародышевых корней, чем у зерновок, убранных в фазе молочной и в начале восковой спелости (табл. 3).
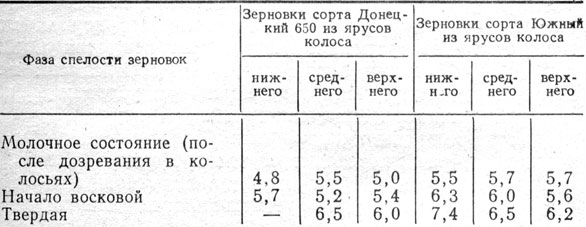
Таблица 3. Число зародышевых корней в зависимости от фаз спелости зерновок ячменя и места их в колосе
Количество зародышевых корней у одного и того же сорта не одинаково по годам. При анализе данных за ряд лет видно, что между числом корней, весом 1000 семян и погодными условиями в период формирования и созревания зерна ячменя существует определенная связь. В 1960 г. во время молочного состояния зерна среднесуточная температура воздуха была 21°С, относительная влажность - 56% и сумма осадков - 4,9 мм, а в 1962 г., когда вес зерна и число корней оказались большими, чем в 1960 г., соответственно 19°С, 68% и 30,5 мм. Чем прохладнее и влажнее погода в период молочного состояния и восковой спелости ячменя, тем крупнее образуются семена, больше закладывается зародышевых корней. Очевидно, при благоприятных погодных условиях полнее протекает процесс формирования зародыша в семени, чем при повышенной температуре воздуха и низкой относительной его влажности.
У крупных семян при прорастании образуется большее число корней, чем у мелких, не выполненных (Синев, 1949; Скрипчинский, 1951; Добрынин, 1952). Чем благоприятнее условия формирования и созревания семян на материнском растении, тем лучше дифференцированы органы зародыша (Морозов, 1949) и большим числом корней они прорастают в почве. Большое значение имеют также условия хранения семян. Фоллстат, Кристенсен (1965) обнаружили, что хранение семян в течение трех месяцев при повышенной температуре (21 °С) не снижало их всхожести, но уменьшало вес корешков в сравнении с хранением при меньшей температуре (18°).
Зародышевые корни играют определенную роль в преобразовании запасных питательных веществ семени в доступную для ростка форму (Малютин, 1959; Мартос, 1959). После прорастания в зародышевых корнях ячменя содержатся стимулирующие рост вещества (Диксон, 1973). Поэтому питательные вещества семени используются для роста побега лишь при условии функционирования зародышевых корней.
Растения из семян, проросших большим числом корешков, скорее переходят к автотрофному питанию. У них впоследствии образуется больше узловых корней, они отличаются лучшей кустистостью и большей продуктивностью (Синев, 1949; Малютин, 1959).
Главный (первый) корень среди остальных зародышевых корней внешне несколько выделяется лишь по толщине, однако роль его в жизни растения весьма велика. Г. М- Добрынин (1969) считает, что без главного корня, развивающегося из его зачатка в зародыше зерновки, растение культурного злака существовать не может. Все эти корни по мере роста углубляются в почву относительно быстро, на каждом из них благодаря ветвлению образуются боковые корни 1-го, 2-го и т. д. порядка, в результате чего формируется зародышевая корневая система. Корни зародышевой системы проникают на глубину 120 см и больше, участвуют в снабжении растения ячменя водой до конца вегетации.
Зародышевые корни отличаются постоянным анатомическим строением, мало зависящим от внешних условий. Колеоптильные корни лишь у основания отличаются от зародышевых большим диаметром, а с глубины 15 - 20 см становятся более похожими на зародышевые. Зародышевые корни, проникая на большую глубину, чем узловые, призваны "черпать" воду из глубоких слоев, в засушливых условиях их функционирование в фазе налива способствует образованию более выполненного зерна.
Зародышевым корням придается особое значение в жизни растений (Чижов, 1931; Жуковский, 1964). Если во время кущения верхний слой почвы быстро высыхает и узловые корни не образуются, как это часто бывает в степной зоне Украины, ячмень развивается на одних зародышевых корнях; продуктивность растений при этом резко снижается.
Через 2 - 3 дня после начала кущения над местом прикрепления первого листа главного побега, из подземного стеблевого узла - узла кущения, образуется первая пара узловых корней. И. В. Красовская (1У52) установила, что эти корни образуются не из улов, а над узлом из меристематической ткани основания вышележащего междоузлия. Можно считать, что именно этим и объясняется то, что узловые корни образуются позже своего побега.
Встречающиеся в литературе высказывания о том, что у ячменя узловые корни образуются позже, чем у овса, требуют уточнения. У овса до появления первого настоящего листа удлиняется первое междоузлие - от узла щитка до узла, к которому прикреплено колеоптиле, и потому часть корней зародышевой системы (колеоптильные) поднимается вверх, ближе к поверхности. У ячменя же эти два узла, а следовательно, и колеоптильные корни остаются на уровне семени. Колеоптильные корни овса в отличие от узловых (появляющихся, как и у ячменя, после начала кущения) проникают вглубь и активно участвуют в питании растения с первых дней вегетации, так же как все корни зародышевой системы.
Размещением в верхнем слое почвы, более богатом микрофлорой и легкодоступными питательными веществами, части активно участвующих в питании растения корней с первых дней вегетации, возможно, и объясняется способность овса неплохо развиваться на почвах с тонким верхним плодородным слоем, преобладающих в северных, более увлажненных районах. Эта особенность овса, как результат его филогенеза, может служить некоторым основанием для выяснения происхождения растения.
Главный стебель образует до 6 - 7 узловых корней и по 2 боковых и более. Так как каждый побег формирует свои корни, число узловых корней находится в прямой зависимости от кустистости. При благоприятных условиях, если растения присыпать почвой, формируются дополнительные корни из вышележащих стеблевых узлов.
Узловые корни более мощные, чем зародышевые. Они также ветвятся, образуя боковые корни 1-го, 2-го и т. д. порядка. Узловые корни распространяются в стороны от стебля, а затем, загибаясь, опускаются вглубь. Распространяются они в основном в пределах пахотного слоя, более плодородном, и потому их роль в питании растения ячменя весьма велика-
Придавая большое значение глубокому размещению корней, нельзя пренебрегать той частью узловых корней, которая размещается в верхней части пахотного слоя. Правда, самый верхний слой почвы бывает в увлажненном состоянии не постоянно, а периодически. Лишь при достаточном его увлажнении деятельность приповерхностной части узловых корней возобновляется, благодаря чему существенным образом улучшается питание растений. Несмотря на это, периодическое высушивание и увлажнение верхнего слоя почвы следует рассматривать не только как явление нежелательное. При чередовании увлажнения и высушивания почва обогащается легкодоступным для растений фосфором. Высушивание верхнего слоя почвы во время вегетации растений и периодическое его быстрое увлажнение во время дождей способствуют заметному увеличению количества фосфорной кислоты, переходящей в почвенный раствор (Францесон, 1949; Лебедянцев, 1960).
Согласно исследованиям В. А. Францесона (1963), в результате высыхания и быстрого увлажнения почвы увеличивается проницаемость оболочек микроорганизмов, вследствие чего вещества, находящиеся в плазме, в составе которых имеется фосфор, получают возможность перейти в почвенный раствор. Наряду с этим увлажнение вызывает также усиление нитрификационных процессов, в результате чего почвенный раствор обогащается азотом. Узловые корни, развивающиеся в верхних слоях почвы, рано высыхающих и лишь периодически увлажняющихся дождями, работают, как отмечают П. В. Морозов (1949), Ф. Балаж (1954), эпизодически от дождя до дождя. Но и при периодическом функционировании в засушливых условиях, как и в районах достаточного увлажнения (Сатарова, 1952), роль их в создании урожая огромна.
Анализ материалов госсортоучастков Днепропетровской области и результатов агротехнических опытов на Эрастовской опытной станции позволяет сделать заключение, что чем больше у растений ячменя сформировалось узловых корней, тем больше зерен было в колосе. В частности, в 1952 г. на вариантах, где среднее число полноценных зерен в колосе главного побега составило 26, количество узловых корней одного растения в фазе выхода в трубку было 8,3, а при пяти узловых корнях в колосе образовалось 20 зерен. В наших опытах на варианте, где среднее число узловых корней у растений ячменя было 6,4, урожай зерна получен 22 ц с 1 га, а при среднем количестве узловых корней 5,1 - 18,2 ц.
Особо благоприятное влияние на число узловых корней и их ветвление оказывают удобрения. В стационарном опыте на Эрастовской опытной станции внесение удобрений под ячмень непосредственно на фоне удобрений, внесенных под предшествующие культуры севооборота, весьма положительно сказывалось на образовании зародышевых, колеоптильных и узловых корней, а также на озерненности колоса и урожае (табл. 4).
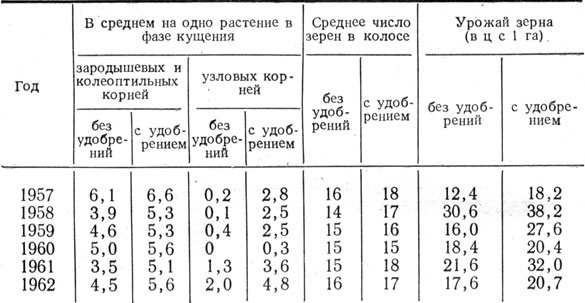
Таблица 4. Влияние удобрений* на укоренение растений ячменя, озерненность колоса и урожай
* (За первую ротацию девятипольного севооборота внесено навоза 20 т на 1 га и N90P150K120, за вторую ротацию - 20 т навоза, N30P30K30 и под ячмень N60P60K60.)
В условиях Эрастовскои опытной станции начало образования узловых корней ячменя приходится, как правило, на вторую декаду мая. Естественно, на формирование узловых корней большое влияние оказывают метеорологические условия этого периода. Так, в 1959 г. при относительно благоприятных метеорологических условиях (среднесуточная температура воздуха 16,5 и С, относительная его влажность 50%, сумма осадков 26,1 мм) за первые десять суток образовалось в среднем на одно растение 3,3 - 3,9 узлового корня; то же наблюдалось и в 1961 - 1964 гг. При более высокой температуре (20,9 и С), низкой относительной влажности воздуха (36%) и малом количестве осадков (8,2 мм) за первые десять суток образовалось в 1958 г. 0,3 - 0,5 узлового корня. Особенно неблагоприятные погодные условия для образования узловых корней сложились в 1966 - 1968 гг. Низкая относительная влажность воздуха и высокая температура почвы сдерживали процесс образования узловых корней. Такую закономерность необходимо принимать' во вни-, мание при выведении новых сортов для степных районов. Чтобы преодолеть пагубное влияние весенней засухи на укоренение ячменя в процессе селекции, следует несколько сместить оптимальную температуру образования узловых корней у растений.
Важная роль корней определяется не только поглощением из почвы воды и минеральных веществ, но и тем, что они участвуют в обмене веществ всего организма. О роли корней ячменя в преобразовании веществ и в процессе синтеза свидетельствуют данные Г. Р. Озолиня, Э. В. Свимпуле (1961), Кейтинг (1963). А. Л. Курсанов (1957, 1960) указывает, что в корне сосредоточена мощная энзиматическая система, действующая как аппарат первичной ассимиляции поглощаемых элементов. Корни являются органами, синтезирующими многие соединения, которые частично используются на месте, частично же увлекаются в надземные органы, дополняя обмен веществ листьев, конуса нарастания и плодов недостающими им соединениями. А. Л. Курсанов (1954) отмечает, что корни участвуют в усвоении растением углекислоты, включая ее и углекислые соли в создание органических соединений. На то, что растения поглощают углерод через корни, указывают также Грехем, Ваунг (1959), Джованоззи-Шерманни (1961). В связи с этим особое значение приобретают агротехнические приемы, способствующие образованию в почве доступной для корней углекислоты в необходимых размерах и создающие оптимальные условия не только для отправления функций поглощения, но и для выполнения корнями тех ответственных превращений, которые составляют часть общего обмена веществ растения.
Нормальная жизнедеятельность растения, обеспечивающая высокую его продуктивность, возможна при непрерывном взаимодействии всех его органов, в том числе и корней- В настоящее время установлено (Колосов и Шалденкова, 1952; Колосов, 1954), что зародышевые колеоптильные и узловые корни, поглощая минеральные вещества и воду, снабжают ими как главный побег, так и боковые побеги растения.
Жизнедеятельность зародышевых корней определяет развитие растения с первых дней вегетации, начиная с прорастания семени. Узловые же корни, которые образуются после начала кущения, участвуют в питании растения из более плодородного пахотного слоя лишь при достаточном его увлажнении. Чем раньше и в большем количестве образуются зародышевые, колеоптильные и узловые корни и чем быстрее они начинают функционировать как органы поглощения, а в связи с этим и участвовать в общем обмене веществ растительного организма, тем более продуктивными бывают растения.
В специальном лабораторно-полевом опыте, где узловые корни первые 10 - 12 дней были изолированы инертной массой (шлаком) и, следовательно, их участие в питании растений начиналось с опозданием продуктивность ячменя снижалась (табл. 5)

Таблица 5. Продуктивность ячменя в зависимости от времени начала участия узловых корней в питании растений на Эрастовской опытной станции (в среднем за 1960-1963 гг.)
При произрастании ячменя в обычной почве, где узловые корни с первых дней развивались в плодородном слое и участвовали в питании ячменя, продуктивность его была заметно выше.
Нами установлено, что повышение плодородия почвы, создаваемого применением удобрений, заметно ускоряет образование узловых корней. В опытах на Урастовской опытной станции в фазе начала кущения растения ячменя на неудобренном фоне не имели узловых корней, а на удобренном имели в среднем по одному корню. В среднем за 2 года в фазе молочного состояния зерна неудобренные растения ячменя имели по 5,1 узлового корня, а удобренные - 6,8.
При нормально развитых зародышевых, колеоптильных и узловых корней почвенное плодородие полнее используется растениями для образования надземной массы и зерна. В районах недостаточного увлажнения при весенней засухе верхний слой почвы часто пересыхает настолько, что формирование узловых корней ячменя ярового задерживается или прекращается совсем. В такие годы продуктивность растений определяется функционированием зародышевых корней. При этом развиваются растения слабее, чем при наличии и узловых. Однако факт плодоношения растений ячменя в годы, когда узловые корни не образуются, свидетельствует о весьма важной роли зародышевых корней и лишний раз опровергает существовавшее в прошлом, а также встречающееся в настоящее время (Комарницкий, Красовская, 1953; Рыжутин, 1970; Рид, Вибе, 1973) мнение о том, что они являются временными и после кущения отмирают или участие этих корней в питании растения резко ослабевает.
Существенное влияние на развитие корней ячменя и их размещение по почвенному профилю оказывают погодные условия - влажность и температура. При избыточном увлажнении почвы корни хлебов, как отмечают И. А. Волков (1951), Я. В. Губанов (1952), Л. В. Мосолова (1956), Т. Ремер (1958), И. А. Стефановский (1965), развиваются слабо, продвижение их вглубь тормозится, они располагаются преимущественно в верхних слоях. При достаточном количестве влаги в глубоких слоях и недостатке ее в верхних корни проникают вглубь (Чижов, 1931; Кружилин, 1936). Но это относится к узловым корням. Зародышевые корни независимо от влажности почвы устремляются вглубь с первых дней вегетации.
По данным П. В. Морозова (1951), Нильсена и др. (1960), с понижением температуры почвы (до 5° C) при достаточном ее увлажнении увеличивалось число и возрастала мощность корней. Поэтому при раннем сроке сева в сравнении с поздним больше образуется разветвленных корней. Влияние температуры на рост корней ячменя можно иллюстрировать данными Эрастовской опытной станции. Растения выращивали в стеклянных трубках диаметром 4,5 см и длиной 140 см, набитых почвой с соблюдением естественного расположения генетических горизонтов. При посеве 22 апреля, когда температура почвы на глубине 5 см в первые восемь дней после посева была в пределах 9 - 10 °С тепла, корни проникали на глубину 10 см, а при посеве 2 июня при температуре 18 °С - на глубину 6 см. В дальнейшем, ко времени образования растениями пятого листа, корни достигали глубины в первом случае 103 см, а во втором - 24 см.
При позднем сроке посева благодаря более высокой температуре образование листьев и рост надземной массы проходили быстрее, чем при раннем, а продвижение корней вглубь, наоборот, резко отставало. Такое же явление наблюдалось при запаздывании с посевом в полевых условиях.
В 1965 г. на Эрастовской опытной станции изучали скорость проникновения корней ячменя сорта Южный вглубь траншейным способом (по методу В. Г. Ротмистрова) в зависимости от крупности высеянных семян и дополнительного увлажнения. Посев был проведен 21 апреля, большинство всходов появилось 5 мая, кущение наступило 28 мая, выход в трубку - 31 мая, колошение - 15 июня, молочное состояние зерна - 21, тестообразное - 28 июня и конец восковой спелости - 5 июля. Увлажнение проводилось после колошения - в период формирования зерна в течение пяти дней вечером из расчета 10 мм ежедневно. В начале вегетации в метровом слое было 128 мм продуктивной влаги, в период колошения и начала формирования зерна на поверхности почвы температура достигала более 44 °С.
Каждый день подсчитывали число корней, которые появлялись на потолке ниш, сделанных в стенке траншеи на разной глубине. Результаты подсчета показали, что на глубине 30 см при высеве более крупных семян первый корень появился на 4-й день после всходов, с каждым днем число их увеличивалось и на 13-й день, когда на этой глубине появление новых корней прекратилось, составило 121. На глубину 120 см проник один корень лишь на 58-й день после всходов, глубже корни не распространялись. При высеве семян с меньшим весом 1000 семян корней образовалось меньше и проникали они не глубже 110 см.
После увлажнения наблюдалось увеличение числа корней глубже 70 см. Например, при высеве более крупных семян на глубине 80 см появилось 116 корней, а после увлажнения - 143, на глубине 90 см соответственно 54 и 65, на глубине 110 - 10 и 26, на глубине 120 см-1 1 и 4 и на глубине 130 см лишь у растений от крупных семян был один корень. Подобные же данные получены и при высеве мелких семян, но корни при этом проникали на глубину не более 110 см. Следовательно, дополнительное увлажнение пахотного слоя в период формирования зерна способствовало улучшению роста корней первичной (зародышевой) системы, которые не только не атрофировались, как предполагалось в прошлом, а продолжали углубляться, и отдельные из них проникали в глубокие слои.
Несколько по-другому влияют условия увлажнения почвы в начале вегетации на создание общей массы корней ячменя. В опытах Эрастовской опытной станции (Г. Г. Дуда, 1958) наличие достаточных запасов влаги в почве в начале вегетации способствовало образованию большей массы корней в метровом слое. В дальнейшем в зависимости от температуры и относительной влажности воздуха мощность надземной массы не всегда соответствовала мощности корней (табл. 6).
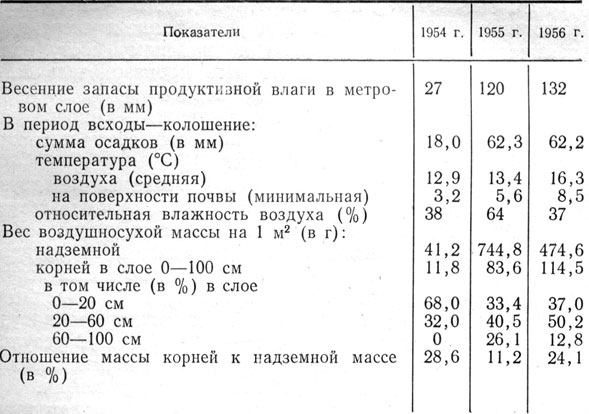
Таблица 6. Влияние погодных условий на развитие корней ячменя
В крайне засушливом 1954 г., когда весной влага была только в верхних слоях и в ничтожно малом количестве, корни образовывались очень слабые, проникали на глубину не более 40 - 50 см, то есть на ту глубину, где имелась влага. Из-за резкого дефицита почвенной влаги вес воздушносухой массы корней ячменя по отношению к весу надземной массы составлял 28,6%. При недостаточном водоснабжении растения ячменя не в состоянии были использовать сформировавшиеся корни для создания соответствующей надземной массы. В благоприятном 1955 г. корни проникли на всю глубину метрового слоя, вес воздушносухой массы корней в этом слое составил 11,2% по отношению к весу надземной массы.
В 1956 г., когда весенние запасы влаги были достаточные, растения образовывали мощную массу корней.
На 1 м2 в слое 0 - 100 см было 114,5 г воздушносухой массы корней. Однако из-за неблагоприятных условий в последующие периоды вегетации (высокая температура и низкая относительная влажность воздуха) надземная масса развивалась значительно слабее, чем в 1955 г. Отношение массы корней к надземной массе было высокое - 24,1%, что указывает на низкую производительность корневых систем. При слабом развитии надземной массы проявлялась тенденция к слабому проникновению корней вглубь. В 1956 г. корней в слое 60 - 100 см было 12,8%, а в 1955 г.- 26,1%. Благоприятные условия в 1955 г.- достаточное увлажнение и умеренная температура - способствовали нормальному росту надземных органов, в результате чего производительность корней была более высокой, чем в 1956 г.
Таким образом, условия увлажнения и температурный режим существенным образом сказываются на характере развития корневой массы.
Стебель ячменя - соломина, состоящая из междоузлий, которых бывает 4 - 7, и узлов. Междоузлия внутри полые, а узлы заполнены. Узлы с зачатками листьев и междоузлий закладываются в начале вегетации, во время кущения. Рост стебля после кущения во время выхода в трубку и колошения осуществляется поочередным удлинением междоузлий. В пределах каждого междоузлия самая молодая ткань в нижней его части. В результате увеличения числа и вытягивания молодых клеток нижней части междоузлий происходит рост стебля. Такой интеркалярный рост стебля возможен благодаря влагалищам листьев, образующим трубку, которая придает прочность побегу с первых дней его формирования.
В начале удлиняется нижнее (первое над узлом кущения) междоузлие, затем - второе и последующие. Междоузлия нижней части стебля возрастно и стадийно старше верхней. У основания стебля междоузлия обычно короче, а к вершине постепенно удлиняются. Однако при благоприятных условиях в период формирования и роста соответствующего междоузлия длина его возрастает. В опытах Всесоюзного научно-исследовательского института кукурузы при улучшении питания растений в начале вегетации при внесении рядкового удобрения второе междоузлие стебля ячменя было длиннее третьего.
В наших опытах на Эрастовской и Одесской опытных станциях выявлено, что чем лучше условия минерального питания в начале роста, тем выше начальные темпы накопления массы и длиннее второе снизу междоузлие стебля. Чем длиннее это междоузлие, тем выше был урожай надземной массы и зерна, и наоборот (рис. 1). Об этом же свидетельствуют и результаты вычисления коэффициента корреляции (табл. 7). Длина междоузлия измерялась по каждому варианту на 40 - 90 растениях. Коэффициент прямолинейной корреляции 0,720 - 0,930 указывает на довольно высокую сопряженность названных показателей. Таким образом, размеры второго междоузлия стебля главного побега ячменя могут свидетельствовать о темпах накопления надземной массы в период его формирования, а при благоприятных условиях в последующем и возможном урожае. В этих опытах коэффициент прямолинейной корреляции между урожаем вегетативной массы и зерна довольно высокий - 0,770 ± 0,130 - 0,980+0,010.
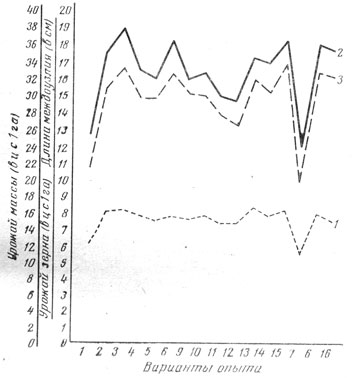
Рис. 1. Взаимосвязь длины второго междоузлия главного стебля (1), урожая надземной массы (2) и зерна (3) ячменя
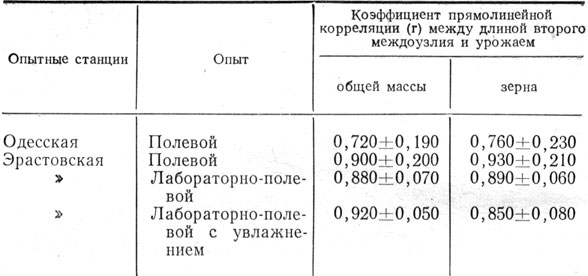
Таблица 7. Коррелятивная связь между длиной второго междоузлия стебля ячменя и урожаем
Одновременно с ростом первого над узлом кущения междоузлия начинается ускоренное развитие зачаточного колоса. Удлинение стебля ячменя в основном заканчивается при полном выколашивании. При благоприятных условиях рост верхнего междоузлия с колосом продолжается и в начале формирования зерен в колосе. Принято считать (Кулешов и Лапцевич, 1951; Пономарев, 1955; Коренев, 1959; Корзун, 1963), что ко времени окончания цветения колоса рост стебля хлебных злаков прекращается.
В опытах на Эрастовской опытной станции при благоприятных условиях увлажнения в период колошения (1961 и 1963 гг.) рост стебля ячменя продолжался в течение трех дней после образования пяточки зерновки, а при недостатке влаги во время колошения (1962 г.) рост стебля прекратился в конце цветения - в начале образования пяточки зерна. В засушливые годы при недостатке почвенной влаги колос ячменя не полностью выходит из пазухи верхнего листа. Высота выноса колоса над пазухой верхнего листа является показателем обеспеченности растений почвенной влагой в период цветения и начала формирования зерновки.
По проводящей системе (сосудисто-волокнистые пучки) стебля перемещается раствор питательных веществ, поступающий от корней, и продукты фотосинтеза к конусу нарастания, а затем к колосу. Во время налива и созревания органические вещества поступают в зерновку в значительной мере также за счет ранее отложенных в стенках стебля. В наших опытах на Эрастовской опытной станции в среднем за 4 года вес надземной массы десяти растений ячменя сорта Южный, начиная от молочного состояния зерновки до твердой спелости, увеличился на 0,15 г, зерна - на 2,23 г, междоузлий стебля уменьшился на 1,92 г, а сорта Донецкий 650 соответственно на 0,77, 2,43 и 1,57 г.
Высокая влажность междоузлий и особенно узлов стебля сохраняется до фазы твердой спелости, причем к концу спелости влажность верхней части стебля оказывается заметно выше нижней. Чем дольше сохраняется высокая влажность верхних узлов и междоузлий стебля, тем полновеснее сформировавшееся к концу вегетации зерно.
Листья на стебле расположены поочередно. Лист состоит из влагалища, листовой пластинки и язычка. Листовое влагалище растений во взрослом состоянии покрывает междоузлие, образуя трубку с несрастающимися по длине краями. Над стеблевым узлом, где лист своей нижней частью прикрепляется к стеблю, образуется кольцеобразное утолщение - листовой узел. Листовой узел защищает нижнюю часть междоузлия, где находится нежная, растущая ткань, от неблагоприятных воздействий и в случае полегания способствует поднятию стебля. В обращенной к почве части листового узла и стеблевого узла начинается усиленное деление клеток, благодаря чему она удлиняется и стебель постепенно поднимается. При окучивании таких узлов можно, как уже отмечалось, вызвать образование корней.
Исследования Г. М. Добрынина (1969) указывают на весьма важную роль листовых влагалищ в формировании побега злакового растения. В период кущения влагалищами листьев создается защита пазушных почек, из которых развиваются боковые побеги. Из влагалищ листьев составляется трубка, внутри которой формируются стебель и соцветие; благодаря этому создаются условия для продвижения побега вверх во время перехода из фазы кущения в фазу выхода в трубку, а затем в фазу колошения.
Таким образом, помимо защиты молодой ткани формирующегося междоузлия стебля и придания ему дополнительной прочности, влагалища листьев вначале способствуют направлению продвижения вверх конуса нарастания стебля, а также определяют направление очередных листьев и зачаточного колоса.
В том месте, где начинается листовая пластинка, влагалище листа заканчивается пленчатым язычком, который переходит в два ушка. Язычок плотно примыкает к стеблю, препятствуя проникновению воды и вредителей внутрь листового влагалища. По длинным клещевидным ушкам, которые, охватывая стебель, заходят концами друг за друга и прижимают к нему влагалище, до колошения можно отличить ячмень от пшеницы и ржи, у которых они небольшие, короткие. После прекращения роста междоузлий стебля влагалища нижних листьев отделяются от него.
Закладка зачаточных листьев (валиков) стебля на конусе нарастания завершается до начала дифференциации зачаточного колоса. Рост же верхних стеблевых листьев проходит одновременно с дифференциацией колоса; это, естественно, сказывается на размере листовой пластинки, так как в первую очередь питательными веществами обеспечиваются развивающиеся генеративные органы. Поэтому длина листовых пластинок, не считая двух нижних, которые формируются еще в зародыше, бывает различная. Размеры листовой пластинки хотя и обусловлены генетически, но являются показателем благоприятности условий среды в период роста листьев соответствующих ярусов (Корнилов, 1968).
Принято считать (Литовченко, 1947; Добрынин, 1952), что первые два листа образуются в основном за счет запасов семени. Однако в наших опытах на Эрастовской опытной станции (обыкновенный малогумусный тяжелосуглинистый чернозем) при внесении удобрений одновременно с посевом пластинки листьев появлялись раньше, быстрее росли и были длиннее, чем без удобрений. Внесение удобрений вблизи семян способствовало удлинению пластинки второго листа с 10,4 до 12,6 - 12,8, третьего с 14,9 до 20,5 - 21,6 см. При недостатке элементов пищи в верхних слоях почвы пластинки второго и третьего листьев ячменя были короче, а внесение удобрений в более глубокие слои обусловило увеличение размера пятого и шестого листьев. Во время выхода в трубку неудобренные растения имели на главном побеге в среднем пять листьев, а удобренные - шесть.
Пластинке листа принадлежит первостепенная роль в формировании габитуса побега. Под действием света в верхушке пластинки первого (всходящего) или последующего листа вырабатываются физиологически активные вещества, обусловливающие развитие всего побега.
В течение вегетативного периода жизни растения подготавливается основа для репродуктивного периода. Чем больше площадь сформировавшихся листьев, тем лучше при благоприятных условиях ассимиляции обеспечиваются продуктами фотосинтеза развивающиеся генеративные органы. Не менее важное значение для генеративного развития имеет фотопериодическая реакция, которая у яровых колосовых начинается при активной физиологической деятельности второго листа. Завершение закладки колосковых бугорков и начало дифференциации элементов будущих цветков обычно совпадают с удлинением первого (над узлом кущения) междоузлия стебля и началом фазы выхода в трубку.
Короткий день и невысокие температуры в период заложения листовых валиков и во время дифференциации колосковых бугорков обусловливают замедление генеративного развития. При замедленном формировании зачатков колоса и благоприятных условиях почвенного питания в последующем в нем образуется больше колосков и зерен. Чем раньше начинает функционировать второй лист после появления всходов, тем продолжительнее период формирования, и потому более развитым при прочих равных условиях бывает колос.
Исследования показали, что соответствующими приемами агротехники можно ускорить формирование второго листа. В наших опытах на Эрастовской опытной станции выявлено, что при высеве тяжеловесных семян ячменя, внесении удобрений в рядки и заделке семян на оптимальную глубину в лучшие сроки появление второго листа растений заметно ускоряется. Помимо этого, более раннее появление листьев связано с удлинением периода их развития при достаточном весеннем увлажнении, быстрым нарастанием ассимилирующей площади и более полным использованием благоприятных условий для накопления продуктов фотосинтеза.
Большое влияние на увеличение площади листовой поверхности оказывают удобрения. На Эрастовской опытной станции у неудобренных растений длина третьего листа в начале колошения была 14,9 см, ширина у основания - 0,71 см, а у удобренных NPK соответственно 20,5 и 0,93 см.
В опытах Измаильской опытной станции (Одесская область, чернозем обыкновенный малогумусный тяжелосуглинистый) Всесоюзного научно-исследовательского института кукурузы ассимилирующая поверхность листьев одного растения ячменя при внесении NPK возросла в фазе трех листочков на 3,7 см2, в фазе выхода в трубку на 18,9 и в фазе колошения на 14,1 см2. Аналогичная закономерность установлена В. А. Дроздовой (1952) и Л. М. Дороховым (1957).
Величина урожая определяется не только общей площадью ассимиляции и продолжительностью ее функционирования, но и продуктивностью фотосинтеза. Это зависит от концентрации хлорофилла в листьях, на что положительно влияет правильное применение удобрений, и, вероятно, также от расположения листьев по отношению к стеблю.
При наклоне листьев от стебля под меньшим углом растения в таком хлебостое для формирования урожая полнее могут использовать солнечную радиацию и другие факторы всеми ярусами листьев, чем при большем наклоне или опущенных листьях. Однако такое предположение пока не подтверждено на практике. Так, почти прямостоячие листья ячменя, свойствены сортам японо-китайского экологического типа, но эти сорта в наших условиях низкоурожайные. Сорта же интенсивного типа Европы имеют поникающие листья (Трофимовская, 1972). Очевидно, в процессе селекции растениям ячменя необходимо придавать способность полнее использовать солнечную радиацию для фотосинтеза при увеличении хорошо освещенной площади всех листьев в посеве.
В последнее время в Австралии (Мельбурнский университет, Ангус, Джонес, 1972) установлено, что у сорта ячменя с прямостоячими листьями выделение углекислоты составило 4,3 г на 1 м2 листовой поверхности в час, а у сорта со свисающими листьями - 3,8 г; листовые влагалища ассимилировали соответственно 13 и 7% всей поглощенной растениями углекислоты. С повышением нормы высева у сорта с прямостоячими листьями отмечено увеличение урожая зерна и сбора сухого вещества, у сорта со свисающими листьями - снижение.
В начале роста листа продукты фотосинтеза используются на формирование его тканей, а после достижения полного размера они постепенно, затем полностью перемещаются в другие органы растения. При старении пластические вещества, а также частично отложенные в пластинке листа перемещаются в более молодые листья и генеративные органы. При нормальных погодных и агротехнических условиях первыми начинают стареть нижние листья, и поэтому пожелтение их в период формирования зерна - явление вполне нормальное.
После усыхания листьев, ко времени молочно-восковой спелости, основными ассимилирующими органами ячменя становятся листовые влагалища и колосья. Последние способны поглощать углекислоту практически до уборки (Шатилов, Ваулин, 1972). Доля колоса в ассимиляции углекислоты в период налива зерна составляет более 50% (Дорохов, Баранина, 1972).
Кущение. При благоприятных условиях увлажнения и питания, невысоких температурах в начале вегетации и оптимальной густоте растения ячменя хорошо кустятся. Однако в засушливых условиях степной зоны значительная часть побегов кущения в процессе вегетации отмирает.
В зависимости от погодных условий выживаемость побегов бывает различной. При благоприятных условиях образуются дополнительные стебли не только от узла кущения, но и от зародышевого или колеоптильного узла - из пазухи колеоптиле. В частности, удобрения, улучшая условия питания в непосредственной близости к прорастающему семени или в зоне распространения молодых корней, способствуют увеличению числа растений с колеоптильными побегами (рис. 2).

Рис. 2. Растение ячменя в период кущения, образовавшее дополнительный стебель от зародышевого узла
На Эрастовской опытной станции в среднем за 2 года при внесении удобрений на глубину 8 - 10 см перед посевом растений со стеблем от зародышевого узла в фазе кущения оказалось 13,1 - 13,2%, а без удобрений - 1,4 - 2,3%. Из-за недостатка влаги и пищи стебли от зародышевого узла полностью погибают. Ни разу не было обнаружено, чтобы такой стебель оказался колосоносным. Однако нет оснований считать, что временное функционирование таких стеблей является вредным для растения в целом.
Образование побегов кущения - это биологическая приспособленность растения произрастать и плодоносить несколькими стеблями. С процессом кущения неразрывно связано образование узловых корней. Образование побегов кущения у ячменя во времени не совпадает с появлением узловых корней: побеги кущения образуются в период появления третьего листа, а корни - при появлении четвертого-пятого листьев (Степанов, 1952). Запаздывание кущения ячменя отодвигает процесс укоренения растений. Это может нанести ущерб растениям, так как, помимо сокращения периода функционирования узловых корней, образование их переносится на период более засушливый, когда верхний слой почвы больше высыхает. Раннее и обильное кущение свидетельствующее о благоприятных условиях для жизни растений, является важной предпосылкой получения растений с хорошей продуктивностью.
Кущению злаков посвящено немало работ, и вместе с тем в литературе нет единого мнения в оценке этого процесса. Широко распространено мнение о том, что существует антагонизм между главным и боковыми побегами, что непродуктивные побеги кущения отвлекают питательные вещества и влагу от главного, снижая его продуктивность. Поэтому в засушливых районах отдают предпочтение одностебельным или малокустящимся растениям, высевают сорта, не склонные к кущению, и применяют приемы, уменьшающие кустистость (Носатовский, 1965).
Нет основания допускать, что в нормальных условиях произрастания растительный организм создает побеги, которые между собой конкурируют, и некоторые из них бесполезны для индивидуума в целом. Боковой побег является составной частью целого организма, и поэтому созданные в корнях, листьях и дополнительных стеблях пластические вещества не остаются изолированными и несомненно идут на пользу всего растения. В наших многолетних опытах выявлено, что степень кустистости растений существенно влияет на их зерновую продуктивность. Наблюдения показали, что чем выше кустистость, тем выше урожай общей массы и зерна растения (табл. 8).
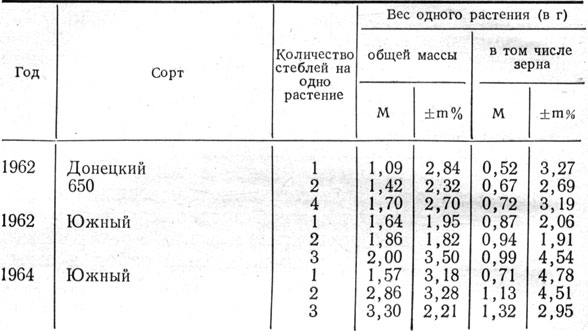
Таблица 8. Продуктивность растений ячменя в зависимости от коэффициента общего кущения
Интенсивность кущения, накопления массы и зерновая продуктивность растений в большой степени зависят от погодных условий в начале вегетации. В степной зоне Украины это главным образом определяется сроком сева (табл. 9). Обусловлено это тем, что при посеве в более поздний срок растения ячменя вегетируют в условиях повышенной температуры и недостатка влаги начиная с первых дней.
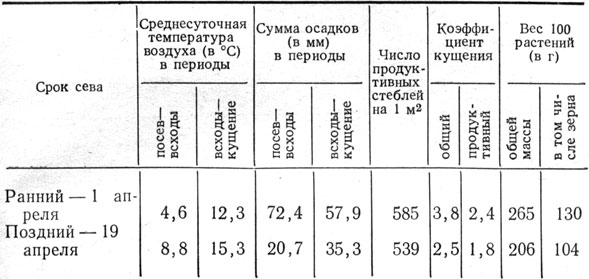
Таблица 9. Коэффициент кущения и продуктивность растений ячменя в зависимости от срока сева (1958 г.)
Основным из факторов, определяющих возможность развития продуктивного побега кущения, является более раннее его образование. Результаты полевых опытов показали, что рано образующиеся побеги кущения, которые ко времени полного кущения имеют не менее трех листьев, могут оказаться продуктивными. Боковые побеги, которые в фазе полного кущения имеют 1 - 2 листа, колоса не образуют. Из 92 растений в момент полного кущения образовалось 37 боковых побегов первого порядка, имевших к тому времени по 3 - 5 листьев, и 109 побегов второго и третьего порядка, имевших по 1 - 2 листа. Ко времени полной спелости из побегов первого порядка образовались озерненные колосья лишь у 14 стеблей, или у 38%, а у побегов второго и третьего порядка не образовалось ни одного озерненного колоса, и задолго до наступления спелости они засохли.
Недоразвитость колоса у побегов кущения второго, третьего и последующих порядков обусловливается в первую очередь тем, что у них все этапы органогенеза оказываются более короткими, чем у главного побега. Чем выше по стеблю расположен побег кущения, то есть чем позже он образуется, тем меньше формируется метамерных органов (Куперман, 1963).
Если рассматривать обильно раскустившееся растение ячменя в целом, то разновозрастные его побеги (главный и боковые), будучи в разном физиологическом состоянии, оказываются под действием одинаковых условий внешней среды. Поэтому у главного и боковых побегов одни и те же этапы органогенеза проходят в разных условиях, в первую очередь температуры. Естественно, что даже нормально развитые побеги, но формирующие свои органы при более высокой температуре, меньших запасах почвенной влаги и питательных веществ, оказываются слаборазвитыми или совсем недоразвитыми. И все же растения с такими недоразвитыми побегами кущения в условиях степной зоны Украины не снижают свою зерновую продуктивность.
В специальном эксперименте Эрастовской опытной станцией выявлено, что при одинаковой продуктивной кустистости увеличение числа непродуктивных стеблей не ведет к снижению сбора зерна. Скорее можно говорить о положительном влиянии общего кущения на зерновую продуктивность (табл. 10).
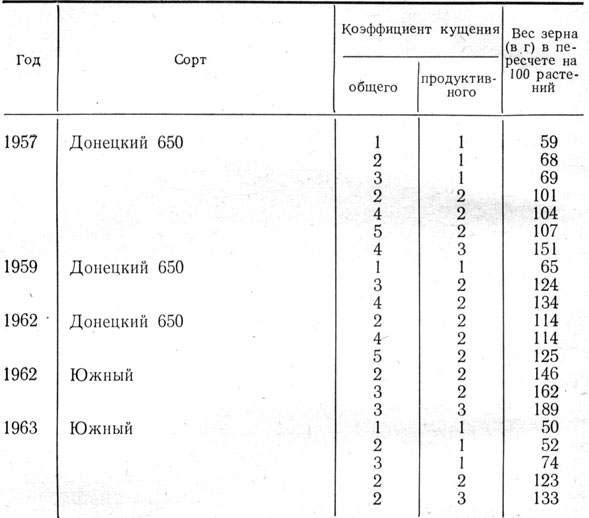
Таблица 10. Вес зерна растении ячменя в зависимости от общего и продуктивного кущения
Увеличение общего кущения в опытах Эрастовской опытной станции никогда не приводило к снижению зерновой продуктивности растений ячменя. Более того, увеличение общей кустистости не влияло отрицательно и
Из данных таблицы видно, что вес зерна главного побега по мере увеличения кустистости несколько возрастал или оставался на том же уровне, что и при меньшей кустистости.
Об участии боковых побегов в питании главного можно судить также по перемещению раствора фуксина кислого, что наблюдалось нами в лабораторных и полевых условиях в 1965 г.
Результаты эксперимента позволяют заключить, что при засушливой погоде, когда нижние листья начинали засыхать, у нормально произрастающего растения (судя по окраске) наблюдалось перемещение веществ из побегов кущения в главный побег ячменя.
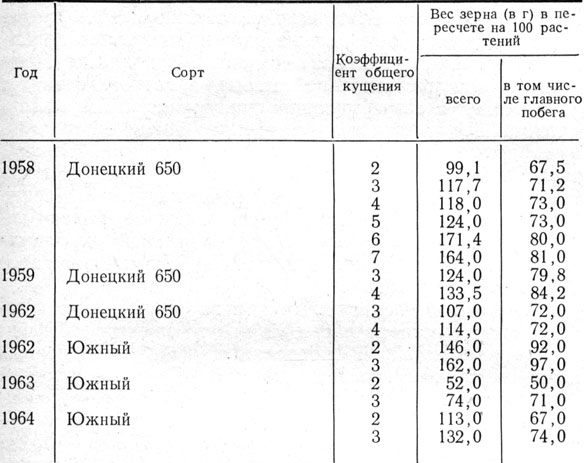
Таблица 11. Урожай зерна ячменя с главного побега в зависимости от общего кущения
Аналогичное отмечали Г. В. Заблуда и М. И. Простева (1956), Квинлен, Сагар (1962), изучавшие передвижение ассимилятов, меченных радиоактивным фосфором или углеродом. В условиях Белоруссии, где побеги кущения благодаря достаточному увлажнению вегетируют до наступления спелости, к ним, как к более молодым органам, перемещаются ассимиляты в первую очередь, что сказывается на ухудшении в результате этого питания главного, более старого побега (Терентьев и др., 1960, 1961, 1962). В условиях же степи Украины, где недоразвитые побеги кущения отмирают за долго до наступления спелости зерна, ассимиляты, образующиеся в тот период, а также мобилизуемые из отложенных в надземных органах, в том числе и в побегах кущения, направляются в формирующиеся органы плодоношения, то есть в первую очередь в колос главного побега.
Способность злаковых растений куститься - это результат длительного процесса приспособления к определенным условиям произрастания. Запасные питательные вещества, накопленные в узлах кущения, обеспечивают ткани растения в период кратковременных, но частых весенних засух, когда фотосинтез снижается до минимума. Наряду с этим способность к усиленному кущению является естественным приспособлением злаков к межвидовой борьбе с сорными растениями.
Хорошо раскустившееся растение затеняет почву и препятствует появлению всходов сорняков. Чем лучше условия питания, тем больше растение образует дополнительных побегов. С этим связано и накопление большего количества надземной массы, а следовательно, и увеличение ассимилирующей поверхности. А максимальной продуктивности растений можно достигнуть при формировании генеративных органов при большом числе крупных листьев (Корнилов, 1957). И действительно, в наших опытах, проводившихся в течение 13 лет, большая общая кустистость растений ячменя сопровождалась более высокой продуктивной кустистостью, густотой колосоносных стеблей, повышением веса 1000 зерен и урожая.
Показателем зависимости уровня урожая зерна ячменя от кустистости растений может служить коэффициент прямолинейной корреляции (г), вычисленный нами по данным госсортоучастков Днепропетровской области за 1944 - 1961 гг. В 37 опытах с сортом Донецкий 650 коэффициент корреляции между общей кустистостью и урожаем зерна составляет от 0,732±0,145 до 0,806+0,109, а между продуктивной кустистостью и урожаем - от 0,841 ±0,045 до 0,965±0,010. О положительном влиянии кущения на урожай зерна указывает также А. А. Наливкин (1947), М. С. Савицкий (1948) и др.
На основании изложенного можно заключить, что в засушливых условиях, где основное значение для урожая ячменя имеют весенние запасы почвенной влаги, наиболее целесообразным является образование боковых побегов в ранние и сжатые сроки, а при наступлении недостатка влаги - сбрасывание недоразвитых и непродуктивных. Раннее интенсивное кущение позволяет молодым растениям быстрее затенять почву в начале вегетации и тем самым уменьшить непроизводительный расход воды на испарение из почвы непосредственно. Исходя из этого, районируемые в степных районах сорта должны отвечать этим требованиям, а соответствующие приемы агротехники - обеспечить усиленное побегообразование с первых дней начала кущения.
Образование побегов и накопление в них пластических веществ способствуют лучшему использованию солнечной энергии для ассимиляции и с помощью узловых корней более полному усвоению растениями питательных веществ и влаги из верхних слоев почвы, откуда она и без того быстро испаряется. Во время дефицита влаги сбрасывание недоразвитых побегов, очевидно, способствует перемещению из них части запасных веществ и ассимилятов в главный побег. В данном случае непродуктивные побеги кущения можно рассматривать как органы, где за счет ранневесенних запасов почвенной влаги накапливается и временно содержится резерв органического вещества, который в нужное время используется растением для создания урожая.
Во влажных районах при обильном кущении запаздывающие в своем развитии побеги кущения могут отвлекать часть пластических веществ, предназначенных для налива зерна колоса главного побега, и затягивать срок уборки. Таким образом, вопрос о полезности кущения для урожая является зональным.
Дифференциация конуса нарастания и закладка репродуктивных органов. Продуктивность колоса ячменя предопределяется в начальных фазах роста и развития растений, задолго до образования видимого простым глазом зачаточного колоса. Более того, И. И. Туманов (1946) считает, что даже величина зерен детерминируется в период дифференциации колоса.
Процесс органообразования, в том числе и образования генеративных органов, находится в тесной зависимости от развития растений. Ф. М. Куперман (1955, 1961) описала семь этапов органогенеза у ячменя. Первый этап характеризуется процессом роста конуса нарастания, второй - незначительным удлинением его и интенсивной дифференциацией зачаточных стеблевых узлов. В этом состоянии растения ячменя могут находиться от фазы проростка до фазы интенсивного кущения. Третий этап органогенеза отличается вытягиванием верхней части конуса нарастания и дифференциацией нижней его части на отдельные сегменты. Четвертый этап характеризуется началом формирования лопастей, из которых образуются колосковые бугорки. Обычно это совпадает с фазой начала выхода в трубку. Лопасти формируются снизу вверх, что обусловливает разновременность появления колосков вдоль колоса и, следовательно, их разнокачественность. Пятый этап органогенеза характеризуется началом формирования цветков в колосках, закладкой колосковых чешуи и протекает в фазе выхода в трубку. На шестом этапе формируются пыльники и завязи, а на седьмом происходит интенсивный рост колосового стержня, покровных тканей, остей и самого стебля.
При благоприятных условиях роста на третьем и четвертом этапах, когда идет дифференциация конуса нарастания на сегменты и закладываются колосковые бугорки, в дальнейшем образуется большее число колосков и более длинный колос. К тому числу бугорков на конусе нарастания (а впоследствии колосков), которое образовалось на третьем и четвертом этапах органогенеза, в последующие периоды вегетации не может прибавиться ни одного, даже при самых благоприятных условиях внешней среды.
Если какая-нибудь часть колоса по какой-либо причине не развивалась в соответствующей ей фазе, то с наступлением развития другого какого-либо признака рост ее прекращается. Так, прекращение развития вершка колосового побега совпадает с моментом появления пыльников в бугорках первых цветков (Сапегин, 1940). После начала появления пыльников все питательные вещества направляются на образование частей цветка, а рост колосового побега и появление новых бугорков - будущих цветков - прекращается.
Таким образом, у ячменя имеются большие резервы увеличения числа колосковых бугорков, так как в верхушке колоса часть их остается всегда недифференцированной на цветки, даже во время формирования половых клеток.
Чтобы увеличить длину продуктивной части будущего колоса, необходимо продлить период образования бугорков - будущих колосков или же создать условия более интенсивной их закладки до появления пыльников. Установлено, что при снижении температуры (Мординова, 1962), затенении в фазе двух-трех листьев (Никитенко, 1951) или сокращении длины дня в период дифференциации конуса нарастания (Образцов, 1971) удлиняется срок образования колосковых бугорков и в колосе ячменя формируется больше колосков. В полевых условиях растения дольше задерживаются на этом этапе органогенеза при умеренно пониженных температурах и облачных днях. При этом рост конуса нарастания и его сегментация растягиваются во времени, в результате чего на нем формируется большее число бугорков, а затем и колосков в колосе.
В горах Северо-Западного Кавказа при выращивании двурядного ярового ячменя в условиях замедленного развития растений и высокой интенсивности фотосинтеза получены шестирядные длинноколосовые и ветвистые формы (Воробьева, 1962). В наших опытах на Эрастовской опытной станции умеренно низкие температуры на четвертом этапе органогенеза обусловили формирование большего колоса с повышенным количеством колосков.
В степной зоне Украины в годы с поздней весной потепление, как правило, наступает быстро, погода бывает безоблачная и третий этап органогенеза проходит скорее. В этих условиях в верхней части конуса нарастания образуется небольшое число колосковых бугорков. Категорическое требование наиболее раннего срока сева ячменя вытекает из необходимости приурочить период формирования зачаточного колоса к наилучшим погодным условиям.
Для нормального развития ячменя на четвертом этапе органогенеза, когда в большей части колосков начинают формироваться половые элементы цветков, весьма важное значение имеет интенсивность освещения (Кудрявцев, 1948; Новиков, 1960). Снижение интенсивности света с начала образования тетрад материнских клеток пыльцы в пыльниках цветков средней части колоса задерживает выколашивание растений и вызывает стерильность большинства колосков (Кудрявцев, 1954; Фесенко, 1963; Гринвалд и др., 1962). Пониженная интенсивность освещения растений в этот период задерживает формирование всех структурных элементов зачаточного колоса и особенно резко сказывается на развитии половых элементов, в результате чего значительная часть даже морфологически вполне развитых цветков оказывается стерильной.
Достаточная влагообеспеченность растений способствует ускорению дифференциации конуса нарастания (Заблуда, 1951). Недостаток влаги во время образования цветков нарушает нормальное течение физиологических процессов прежде всего в молодых колосках, расположенных в верхней части колоса, в результате чего они остаются бесплодными (Заблуда, 1938, Прилл-витц, 1964).
Ячмень очень чувствителен к недостатку воды, особенно когда формируется пыльца (Аникиев и Горощен-ко, 1950; Сказкин и Фонталина, 1951; Яковска, 1963; Рубинчикова, 1965). Пыльца, как наиболее чувствительный элемент цветка растения, испытывает на себе последействие засухи, в результате чего большое количество ее оказывается стерильной. Это неоднократно наблюдалось и нами в полевых опытах на Эрастовской опытной станции. Так, в 1965 г., когда во время формирования колоса почти не было осадков (в мае выпало 18,3 мм и в июне 23,8 мм, из них полезные лишь 14 мая 11 мм и 12 июня - 6,1 мм), а исходные запасы почвенной влаги были ограничены, наблюдалась массовая стерильность верхних колосков, в ряде случаев в колосе было более половины стерильных цветков - завязь развилась, но пыльцы не оказалось.
Решающее значение для формирования основы генеративной части растений имеют благоприятные условия питания в начале вегетации (Вейдеман, 1927; Дикусар, 1939; Сабинин, 1940; Бущуева, 1955). При удлинении третьего этапа органогенеза и обильном питании Ф. М. Куперман (1950) наблюдала образование многоцветковости в колоске ячменя. Поэтому существенное влияние на образование зачаточного колоса, а затем и на формирование его продуктивности оказывает степень плодородия верхнего слоя почвы, определяющего уровень питания молодых растений.
На Эрастовской опытной станции в период от появления второго листа до полного кущения изучали динамику роста конуса нарастания стебля ячменя в зависимости от сроков и способов внесения минеральных удобрений. Из данных измерения длины конуса нарастания растений (табл. 12 и рис. 3, 4) видно, что внесение полного минерального удобрения заметно ускоряло темпы роста конуса нарастания стебля и начало его дифференциации. Наибольшие темпы роста конуса нарастания наблюдались при локальном внесении NPK на глубину 5 см; отмечается более ускоренный рост стебля ячменя благодаря удлинению первого надземного междоузлия. Аналогичные результаты получены и в полевом опыте.
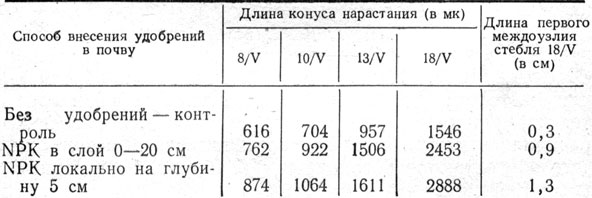
Таблица 12. Влияние минеральных удобрений на динамику роста конуса нарастания стебля ячменя в зависимости от способа внесения их в почву осенью
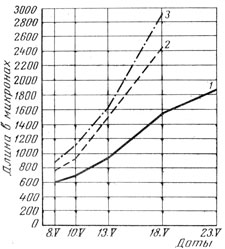
Рис. 3. Динамика роста конуса нарастения главного стебля ячменя в зависимости от способа внесения удобрений в почву осенью: 1 - без удобрений; 2 - NPK в слой 0-20 см; 3 - NPK сосредоточенно на глубину 5 см
Положительное влияние удобрений на рост конуса нарастания подтверждают также данные наблюдений, полученные в опыте с внесением полной дозы NPK и Р в рядки при посеве ячменя.
Заслуживает особого внимания тот факт, что внесение одного фосфора в рядки влияло на скорость роста конуса нарастания почти так же, как и полное минеральное удобрение. Очевидно, сказывается известное физиологическое значение фосфора в жизни растений, в частности связанное с процессами дыхания и фотосинтеза, обусловливающими накопление больших количеств энергии. Возможно, имеет значение его роль в обмене веществ, в первую очередь то, что фосфор входит в состав нуклеопротеидов, высокое содержание которых свойственно меристематическим тканям растения (Рубин, 1961). Это положение подтверждается тем, что растения, как установлено многочисленными опытами, нуждаются в фосфорном питании с первых дней своей жизни (Шестаков, 1954).
Таким образом, условия минерального питания оказывают существенное влияние на закладку, дифференцирование и рост репродуктивных органов, что в конечном итоге выражается общим количеством цветков.
Результаты полевых и лабораторно-полевых опытов позволяют заключить, что применение удобрений, и особенно внесение их в рядки, оказывает благоприятное влияние не только на закладку генеративных органов, но и на увеличение количества зерен в колосе. Есть основание считать, что на обыкновенном черноземе наиболее важным элементом в составе полного минерального удобрения, влияющим на продуктивность колоса ячменя, является фосфор.
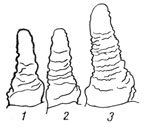
Рис. 4. Динамика дифференциации конуса нарастания главного стебля ячменя в зависимости от способа внесения удобрений в почву осенью: 1 - без удобрений; 2 - NPK. в слой 0-20 см; 3 - NPK сосредоточенно на глубину 5 см
Рис. 4. Динамика дифференциации конуса нарастания главного стебля ячменя в зависимости от способа внесения удобрений в почву осенью: 1 - без удобрений; 2 - NPK. в слой 0-20 см; 3 - NPK сосредоточенно на глубину 5 см
Рис. 4. Динамика дифференциации конуса нарастания главного стебля ячменя в зависимости от способа внесения удобрений в почву осенью: 1 - без удобрений; 2 - NPK. в слой 0-20 см; 3 - NPK сосредоточенно на глубину 5 см
Рис. 4. Динамика дифференциации конуса нарастания главного стебля ячменя в зависимости от способа внесения удобрений в почву осенью: 1 - без удобрений; 2 - NPK. в слой 0-20 см; 3 - NPK сосредоточенно на глубину 5 см
Колос - соцветие ячменя - состоит из стержня и прижатых к нему колосков (цветков). Колосовой стержень представляет собой ступенчатую зигзагообразную линию, как бы составленную из отдельных члеников. На выступах колосового стержня помещаются по три одноцветковых колоска. Если все колоски, сидящие на выступе колосового стержня, плодущие и образуют зерновки, колос шестирядный или многорядный (подвид Н. vulgare L.); если же зерновка образуется лишь в среднем колоске - колос двухрядный (подвид Н. disti-chum L.). Бывают и промежуточные формы, у которых количество плодущих колосков неопределенное (подвид Н. intermedium Vav. et Orl.).
В каждом развитом колоске, помимо двух узких колосковых чешуи, имеется один цветок, состоящий из генеративных органов (одна завязь, три тычинки), лодикулы и двух цветковых пленок, которые у пленчатых ячменей срастаются с зерновкой. Наружная цветочная пленка у остистых форм образует ость - гладкую или зазубренную, а у некоторых ячменей вместо ости образуется трехлопастный придаток (фурка). Ости участвуют в процессе зернообразования; при суховеях они способствуют усилению притока влаги в колос.
У ячменя в отличие от других колосовых на вершине колоса имеются недоразвитые отмершие колоски. Это свидетельствует о том, что при соответствующих благоприятных условиях в период развития колоса на нем может сформироваться значительно большее число колосков.
Цветение ячменя, как открытое, так и закрытое, в основном совпадает с полным выколашиванием. Вместе с тем, по мнению П. Ф. Гаркавого (1949), Г. Ф. Никитенко (1950), А. И. Руденко (1950), М. П. Манжос (1962), опыление ячменя, особенно при засухе, происходит, когда колос еще не вышел из влагалища верхнего листа. При благоприятных же условиях на плодородных почвах, как установил А. М. Мироненко (1953) на Синельннковской селекционно-опытной станции, растения ячменя выколашиваются дружно и цветение наступает после выхода колоса из листового влагалища.
Общая продолжительность периода колошения зависит как от погодных условий, так и от особенностей сорта. От начала колошения сорта Донецкий 650 до полного его завершения в 1961 г. прошли 21 сутки и в 1962 г. 16 суток, а сорта Южный соответственно 22 и 18 суток. Некоторое увеличение продолжительности периода колошения растений сорта Южный объясняется тем, что у него побеги кущения, за счет которых и растягивается выколашивание, развиты слабее, чем у Донецкого 650.
Сортовые особенности, проявляющиеся у растений сорта Южный в более сжатых сроках массового колошения главного побега и задержке выколашивания побегов кущения, в сравнении с сортом Донецкий 650 могут быть объяснены своеобразием создающегося микроклимата в стеблестое. Благодаря обильному кущению растения сорта Донецкий 650 лучше прикрывают поверхность почвы, чем сорта Южный, что вызывает меньший прогрев ее. Так, в 1961 г. в течение четырех дней (12 - 16 июня) средняя температура на поверхности почвы в посевах ячменя сорта Донецкий 650 в 13 часов была 29,6 °С, а сорта Южный - 30,4° С; в 1962 г. (5 - 8 июня) соответственно 17,1 и 19,9°С.
Заслуживает внимания тот факт, что в неблагоприятных условиях 1962 г. период выколашивания был короче на 4 - 5 дней, чем в 1961 г., когда за 10 дней (5 дней до начала колошения и 5 дней после него) выпало 30,4 мм осадков, среднесуточная температура воздуха была на 3,1° С меньше, а относительная влажность воздуха на 23,8% выше, чем в 1962 г. Цветение колосков средней части колоса в благоприятные годы заканчивалось, когда он выходил на 3/4 своей величины из влагалища. Аналогичное явление наблюдали И. М. Мантурова, П. Д. Вершинина (1959) в условиях Крайнего Севера. Следует, однако, подчеркнуть, что в условиях степи бывают годы, когда из-за недостатка влаги колос вообще не выходит полностью из листового влагалища.
Таким образом, по нашим наблюдениям, выход колоса из листового влагалища и продолжительность роста верхнего междоузлия после этого зависят от погодных условий и потому в отдельные годы проявляются по-разному. Следовательно, определение времени цветения по состоянию выхода колоса из влагалища не надежно. Вместе с тем продуктивность колоса определяется сроком его появления из влагалища верхнего листа. При более раннем появлении колоса продуктивность его выше, чем при позднем (табл. 13).
Меньшая продуктивность позже появившихся колосьев в значительной степени определяется и тем, что они состоят в основном из колосьев побегов кущения.
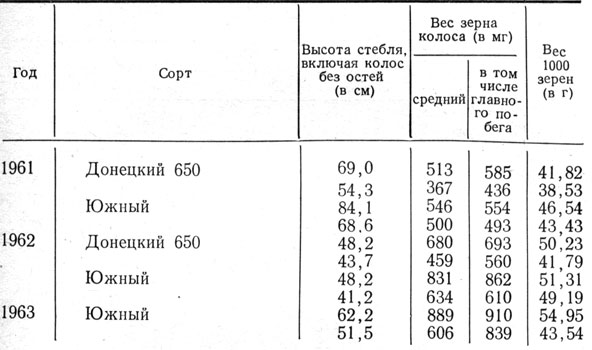
Таблица 13. Продуктивность колоса ячменя в зависимости от срока его появления из влагалища листа
Примечание. Первая строка-колос появился в первый день колошения; вторая - в конце фазы.
Раздельный учет числа цветков, зерен, веса зерна колоса и веса 1000 зерен (табл. 14) показал заметное отставание колосьев побегов кущения, особенно позже появившихся, от колосьев главного побега.
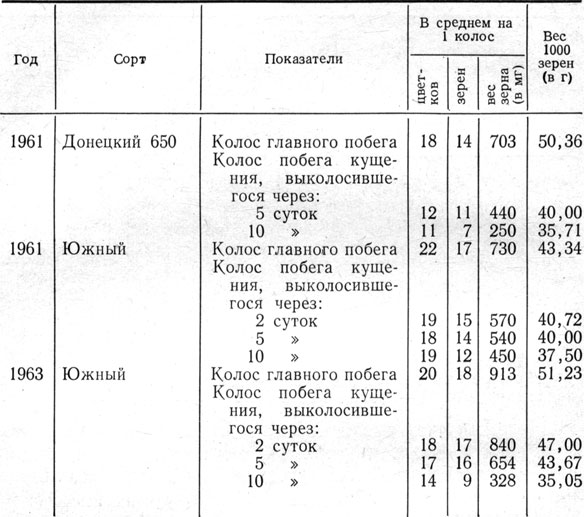
Таблица 14. Сравнительная продуктивность колоса главного побега и побегов кущения разных сроков выколашивания
Тяжеловесность зерен и их количество в колосе в значительной степени обусловлены погодными условиями в период формирования зерна и общим состоянием развития растений к тому времени. В связи с этим сроки появления колосьев предопределяют более полный налив зерна или же ограничивают эту возможность.
Зернообразование. Полученный нами шестилетний экспериментальный материал свидетельствует о том, что в начале формирования зерновки влажность ее наиболее высокая и по мере накопления сухого вещества и созревания постепенно снижается (табл. 15, рис. 5). Ко времени окончания роста зерновки в длину, что отмечалось через 10 - 11 дней после цветения, влажность зерна была 68,5 - 70,2%.
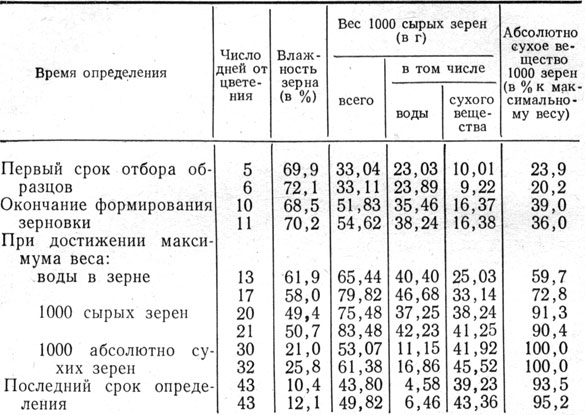
Таблица 15. Накопление сухого вещества и динамика содержания воды в зерне ячменя в процессе зернообразования
Примечание. Верхняя строка - данные по сорту Донецкий 650, нижняя - по сорту Южный.
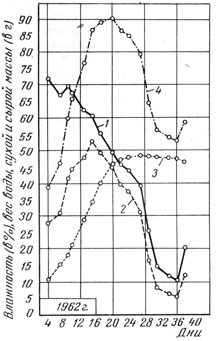
Рис. 5. Изменение содержания воды и сухого вещества в зерне ячменя сорта Южный в период зернообразования: 1 - влажность (в %); 2 - вес воды в 1000 зерен (в г). 3 - вес абсолютно сухого вещества 1000 зерен (в г); 4 - вес 1000 сырых зерен (в г)
После завершения фазы формирования зерновки в течение 2 - 5 дней у сорта Донецкий 650 и в течение 5 - 8 дней у сорта Южный наблюдалось увеличение веса воды в сыром зерне, а затем неуклонное его уменьшение. Когда вес воды в 1000 сырых зерновках достиг своего максимума, влажность зерна в сравнении с предыдущим определением несколько снизилась. Снижение влажности зерна при одновременном увеличении веса воды в 1000 сырых зерновках свидетельствует о весьма интенсивном накоплении сухого вещества в этот период.
Наступление максимального веса 1000 сырых зерновок по времени не совпадает с достижением их наибольшего веса абсолютно сухого вещества. Наибольший вес 1000 сырых зерен наступал в зависимости от года на 5 - 14 дней раньше, чем наибольший вес сухого вещества зерна.
К моменту достижения наибольшего веса абсолютно сухого вещества зерна влажность его в среднем составляла 21 - 25,8%. Так, у ячменя сорта Донецкий 650 через 25 - 34 дня после цветения влажность зерна была в 1957 - 1960 гг. 18,2 - 20,8%, в 1961 - 1962 гг.-23,1 - 23,4%, а у сорта Южный через 30 - 36 дней в 1960, 1961, 1963 и 1965 гг.- 20,1 - 24,9%, в 1962 г. через 27 дней - 39,1%. В условиях Канады (Мак Грегор и др., 1971) накопление сухого вещества в зерновке ячменя отмечалось до тех пор, пока влажность его не снизилась до 27%. Это не согласуется с имеющимися указаниями на то, что поступление веществ в зерновку ячменя при снижении ее влажности до 32 - 42% прекращается (Харлан, Поп, 1923; Медведев, 1937; Гесслейн, 1959; Старжицки, Шепенец, 1960).
Максимальный вес абсолютно сухого вещества зерна наступает в конце восковой спелости. Как правило, наиболее высокий вес 1000 зерен наблюдается в годы, когда во время интенсивного налива температура невысокая и период зернообразования более продолжительный. Так, в 1958 и 1962 гг., когда в указанный период выпало 54,1 - 67,7 мм осадков, температура в 13 часов на поверхности почвы в стеблестое сорта Донецкий 650 была 28,4 - 28,9 °С, вес 1000 зерен был 44,27 - 46,17 г, а в 1960 г. при температуре 39,6 °С и сумме осадков 11,6 мм - 33,45 г.
Процесс налива связан с быстрой мобилизацией и передвижением в колос из вегетативных органов питательных веществ, для чего необходимо достаточное водоснабжение (Максимов, 1952). До молочного состояния зерна в растении протекают процессы синтеза, при которых усваивается большая часть минеральных веществ, необходимых для процессов обмена (Богданов, 1953). После этой фазы преобладают процессы гидролиза и часть минеральных веществ переносится в зерно. Соединения азота в зерновку поступают из запасов, накопленных в растении до цветения (Недокучаев, 1922; Тулайков и Писаревский, 1927; Крокер и Бартон, 1955; Таранов, 1962; Ниле и др., 1963). При недостатке воды и ускоренном созревании миграция минеральных веществ в формирующиеся репродуктивные органы ускоряется (Коик и др., 1962).
Наши исследования показали, что в стеблевых узлах содержится много влаги'вплоть до наступления твердой спелости. Например, в среднем за 1959 - 1962 гг. у растений сорта Донецкий 650 во время молочного состояния зерна во всех стеблевых узлах содержалось 59,6% воды, в том числе в первом от поверхности земли узле 52,5%, в пятом 66,1%; во время твердой спелости влажность узлов была соответственно 52,4; 40,6 и 61,4%. По мере созревания влажность стеблевых узлов несколько снижалась, но это относится главным образом к нижним. В верхних же узлах влажность оставалась довольно высокой также при твердой спелости.
Высокая влажность сохраняется долго, до полного созревания, не только в стеблевых узлах, но и в междоузлиях стебля и заметно уменьшается в листьях и в колосовом стержне. В среднем за 4 года у растений сорта Донецкий 650 при твердой спелости влажность зерна была 14,2%, всех междоузлий - 42%, в том числе нижнего 40,4 и верхнего 37,3%, а листьев с листовыми влагалищами соответственно 16,6; 7,9 и 18,7%.
Заслуживает внимания, что во время твердой спелости влажность верхнего междоузлия стебля в отличие от узлов меньше, чем нижнего, что наблюдали также Старжицки и Шепенец (1960). Очевидно, в период зернообразования из верхних междоузлий влага расходуется интенсивнее, чем из нижних. Влажность верхнего листа во время восковой и твердой спелости всегда выше влажности колосового стержня и зерна, что наблюдали также Н. Н. Овчинников, Н. М. Шиханова (1965). Это в определенной мере свидетельствует о том, что листья верхнего яруса играют большую роль в снабжении репродуктивных органов ассимилятами в период налива.
Интересно, что в пределах междоузлий стебля также существуют различия в содержании влаги. В нижней половине пятого междоузлия влажность составляла во время колошения 82,1%, во время молочного состояния зерна - 52,8 и при твердой спелости - 49,2%, а в верхней соответственно 75,5; 51,3 и 38,4%. Об этом свидетельствуют также А. И. Меркис, Л. Л. Приалгаускайте (1963) в отношении активности ферментов и Торн (1959) в отношении интенсивности дыхания. Очевидно, объясняется это тем, что в силу интеркалярного роста стебля наиболее молодая ткань его находится в нижней половине междоузлия; по мере удаления от основания стеблевого узла ткань междоузлия является возрастно более старой.
Насколько важным для налива зерна является достаточная оводненность вегетативных органов, можно судить по опыту 1960 г., когда во время наступления твердой спелости влажность стеблевых узлов, междоузлий стебля, листьев с листовыми влагалищами и стержня колоса сорта Донецкий 650 была наименьшей за все годы изучения (соответственно 46,0; 29,I; II и 10,2%), вес 1000 зерен был самый низкий - 33,45 г и процесс зернообразования закончился за короткий период - за 27 дней.
Принято считать, что от начала молочной спелости до твердой средний сухой вес растения не увеличивается, несмотря на быстрое увеличение веса зерна. Следовательно, зерно образуется за счет вегетативных органов. Действительно, за период от колошения до твердой спелости отмечается существенное перемещение сухого вещества в отдельных органах надземной массы. В 1960 г. прирост веса сухого вещества зерна десяти колосьев от фазы молочного состояния до твердой спелости (от 4,65 до 5,52 г) составил 0,87 г, то есть почти столько же, сколько и убыль сухой массы междоузлий и листьев,- 0,88 г (4,74 и 3,86 г). Во все остальные годы, когда влажность вегетативных органов была выше, чем в 1960 г., прирост зерна за названный период заметно превышал убыль в весе сухой массы междоузлий и листьев. Например, в 1959 г. за период от фазы молочного состояния зерна до твердой спелости прирост сухого вещества зерна десяти колосьев был 2,81 г, а убыль веса междоузлий и листьев - 1,11 г, в 1961 г. соответственно 2,26 и 1,85 г, в 1962 г.- 3,80 и 2,42 г.
У неудобренных растений вес сухой массы колосьев во время колошения составил 19,7% веса всей надземной массы колосоносных стеблей, а во время твердой спелости - 58,8%. Вместе с тем вес стеблей с листовыми влагалищами уменьшался с 53,3 до 30,4%, а листовых пластинок - с 27 до 10,8%. У удобренных растений наблюдалась такая же закономерность, хотя в отличие от неудобренных они в одинаковых погодных условиях оказались способными после цветения накапливать органическую массу в заметных размерах. Следовательно, зерно у неудобренных растений образуется преимущественно за счет пластических веществ, накопленных в основном ко времени полного колошения, а у удобренных - за счет накопленных и позже.
Таким образом, налив зерна происходит как за счет притока готовых (запасных) пластических веществ из вегетативных органов, так и благодаря текущей ассимиляционной деятельности зеленых органов, на что указывал также ряд авторов (Терентьев и др., 1961; Джонард, 1962; Бирекка и др., 1964; Дилз, 1964).
Образующиеся в период зернообразования углеводы благодаря фотосинтетической деятельности полностью откладываются в семенах, так как формирование вегетативных органов к тому времени закончено. Однако размеры прироста веса зерновки зависят от развития растений и оводненности междоузлий стебля. Например, в загущенных посевах, где рост вегетативной массы ограничен, образование массы зерна в значительной степени обеспечивается за счет запасов, ранее накопленных в вегетативных органах. В разреженных посевах, где растения имеют мощно развитые вегетативные органы, налив зерна длительное время осуществляется за счет вновь образующихся в процессе фотосинтеза ассимилятов. В изреженных посевах ко времени уборки в вегетативных органах содержатся еще большие запасы неиспользованных на налив зерна питательных веществ (Кравцова, 1957).
Чтобы выявить, когда прекращается поступление веществ из вегетативных органов в колос, в конце созревания на Эрастовской опытной станции проводили опыты с помощью красителя. Двадцать одинаково развитых колосьев главных побегов вместе с верхним междоузлием срезали над верхним стеблевым узлом под водой и немедленно погружали в колбу, установленную среди посева с 1%-ным раствором фуксина кислого. В каждой фазе спелости отобранные колосья погружали в раствор в 9 - 10 часов утра и оставляли там в течение четырех часов.
Наблюдения в 1965 г. показали, что в конце восковой спелости окрашивание всех частей колоса, как и в предыдущей фазе, было вполне заметно. Цветковые чешуи окрашивались вдоль прожилок отдельными линиями. Это указывает на то, что физиологическая связь зерновки с материнским растением еще не прервана. Очевидно, проводящие пути стеблей сохраняют способность перемещать влагу и пластические вещества в репродуктивные органы до твердой спелости.
При благоприятных погодных и агротехнических условиях передвижение пластических веществ в колос из листовых пластинок, листовых влагалищ и стеблей внешне проявляется в постепенном отмирании листьев, начиная с нижних. Такая очередность увядания листьев в период зернообразования способствует лучшему наливу зерна. При этом зерно воспринимает полностью весь заготовленный для него запас пластических веществ, и лишь после совершенной выполненности зерна налив заканчивается естественным образом.
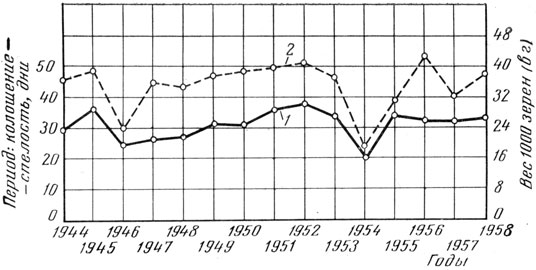
Рис. 6. Зависимость веса 1000 зерен ячменя от продолжительности периода колошение - твердая спелость: 1 - вес 1000 зерен (в г); 2 - продолжительность периода колошение - твердая спелость (в днях)
В годы с сильной воздушной засухой во время налива и созревания ускоренное отмирание листьев происходит также сверху стебля вниз и период зернообразования резко сокращается. В результате засыхания ножки, соединяющей зерно с материнским растением, получается преждевременное физиологическое разобщение, и пластический материал остается в вегетативных органах. Такие неблагоприятные условия были в 1953 и 1964 гг., когда вес 1000 зерен был наименьший - 31,76 и 25,88 г.
Сокращение периода зернообразования в засушливые годы в значительной мере зависит от температуры. При высокой температуре ускоряются все химические процессы, протекающие в зерне, в результате чего зерно быстрее проходит все фазы спелости. Вместе с тем перемещение пластических веществ из вегетативных органов в зерновку более полно происходит при некотором удлинении процесса. Как правило, удлинение периода формирования, налива и созревания зерна указывает на то, что погодные условия благоприятствуют этому процессу-В связи с этим повышенный вес 1000 зерен, как правило, наблюдается в те годы, когда период колошение - твердая спелость бывает более продолжительным, что видно из данных Синельниковского госсортоучастка за 1944 - 1958 гг. Коэффициент корреляции равен 0,744±0,103 (рис. 6). Это подтверждается также материалами госсортоучастков Днепропетровской области в среднем за 17 лет (табл. 16).
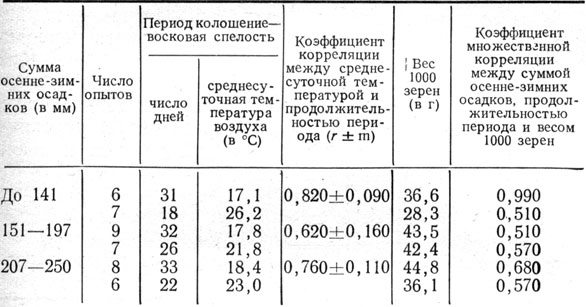
Таблица 16. Вес 1000 зерен ячменя в зависимости от продолжительности периода колошение - восковая спелость и среднесуточной температуры
Естественно, что продолжительность периода налива тесно связана не только с погодными условиями, но и с особенностями растительного организма, которые, в свою очередь, определяются сортовыми различиями и приемами выращивания. При наличии влаги в почве и нормальном перемещении ее в растении поступление пластических веществ из вегетативной массы в зерновку, очевидно, возможно и в последние дни зернообразования, когда зерно уже вступает в фазу твердой спелости. При довольно высокой влажности междоузлий стебля во время восковой спелости в 1959, 1961 и 1965 гг. в отличие от 1960 г. продолжалось накопление сухого вещества в зерне и при влажности его ниже 20% до наступления твердой спелости. В 1960 г. при наиболее низкой влажности верхних междоузлий и листьев (43 и 18,8%) во время восковой спелости поступление пластических веществ в зерновку завершалось при ее влажности 30 - 40%. Осадки, выпавшие позже, когда проводящие пути, по-видимому, уже были нарушены, не способствовали продолжению поступления органического вещества в зерновку.
Таким образом, результаты наших исследований й данные других авторов свидетельствуют о том, что при умеренно теплой и достаточно влажной погоде во время налива и созревания период зернообразования удлиняется. При этом прирост веса зерна продолжается до твердой спелости и вес 1000 зерен бывает выше. Если же в период налива и созревания бывает слишком высокая температура почвы и воздуха и низкая относительная влажность воздуха, перемещение пластического материала из листьев и стебля в зерно преждевременно приостанавливается из-за нарушения нормального передвижения воды в растении. В результате этого вес 1000 зерен снижается.
Поступление пластических веществ из вегетативных органов в зерновку ячменя при определенных условиях на Эрастовской опытной станции наблюдалось и после скашивания в ранние фазы (табл. 17).

Таблица 17. Вес 1000 зерен ячменя в конце восковой - начале твердой спелости при немедленном отчленении от колоса и дозревании в валках
Вместе с тем в ряде работ указывается, что после скашивания ячменя перемещение веществ из вегетативной массы в зерновку прекращается. Разноречивые суждения по этому вопросу объясняются тем, что возможность перемещения пластических веществ в зерновку после скашивания определяется рядом условий: мощностью развития вегетативной массы и состоянием спелости зерна в момент скашивания, толщиной и плотностью валка, погодными условиями в первое время после скашивания. При быстром высыхании скошенной массы в валках, что наблюдается в знойную пору, функционирование проводящих тканей нарушается, и даже при влажности зерна выше 40% приток пластических веществ к ним исключен.
Если растения находятся в состоянии, близком к твердой спелости, когда прироста веса зерна на корню уже не бывает, то тем более его не будет после скашивания в такое время. При благоприятных же погодных условиях, когда скошенная масса в валках подсыхает постепенно, приток питательных веществ в зерновки, особенно отставшие в своем развитии (в колосьях побегов кущения, в колосках нижнего и верхнего ярусов колоса), некоторое время продолжается. Обусловлено это тем, что перемещение веществ в зерновку из стебля и листьев по инерции продолжается и после скашивания. Наряду с этим снабжение зерновок питательными веществами благодаря фотосинтетической деятельности колоса (Батроус, 1959; Торн, 1965; Бирекка и др., 1964) нескоро прекращается после скашивания, если погодные условия не вызывают быстрого высыхания вегетативной массы. Более того, В. Г. и О. Г. Александровы (1940) выявили наличие хорошо развитой гидроцитной ткани в колосковой чашуе и окремнелых бугорков в эпидермисе, способствующих накоплению и распределению воды, необходимой для ассимиляционного процесса и для развития зерна. Это способствует поступлению воды в колоски и зерновку, помимо подачи ее корневой системой.
Учитывая вышеизложенное, можно предположить, что в отдельных случаях при определенных погодных условиях и сохранении зеленого пигмента в колосьях возможно увеличение веса зерновок при частичном высыхании скошенных стеблей. При этом не следует исключать роль остей в наливе зерна (Апель, 1966; Сагир и др., 1968; Шаллер, 1972).
Благодаря продолжению после скашивания фотосинтетической деятельности вегетативных органов растения и составных частей колоса вес 1000 зерен в валках возрастает преимущественно за счет уменьшения числа мелких, недоразвитых. В многочисленных работах (Коданев, 1956; Бялый, Никифоров, 1959; Суднов, 1959) отмечается, что во многих случаях после наступления наибольшего веса зерна при созревании вскоре начинается его уменьшение.
Снижение веса зерна ячменя при оставлении растений на корню после достижения конца восковой спелости установлено и в наших опытах (табл. 18).
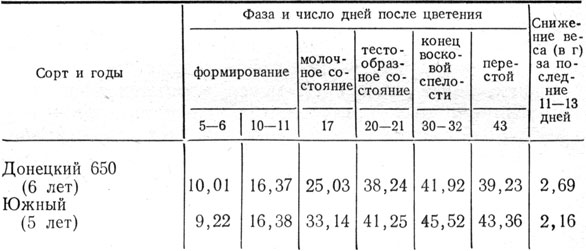
Таблица 18. Динамика веса 1000 зерен ячменя во время зернообразования
Примечание. Показан средний вес абсолютно сухих 1000 зерновок в конце каждой фазы.
После наступления максимума веса 1000 зерен при оставлении на корню в течение 11 - 13 дней вес сухого вещества 1000 зерен уменьшился у сорта Донецкий 650 в среднем за 6 лет на 2,69 г, а у сорта Южный в среднем за 5 лет на 2,16 г. В годы, когда полезные осадки сочетались со сравнительно высокой относительной влажностью воздуха, потери были большие. Например, в 1960 г., когда во время перестоя выпало 35,2 мм осадков и относительная влажность воздуха была 54,3%, потери сухого вещества зерна сорта Донецкий 650 за 9 дней составили 11,3%. Заслуживает особого внимания тот факт, что в первые семь дней, когда зерно было в фазе твердой спелости, среднесуточные потери сухого вещества значительно меньше, чем в последние дни перестоя (табл. 19).
Такое же явление наблюдал А. И. Лебедик (1957) в опытах с озимой пшеницей- По мнению ряда авторов, потери в весе зерна после достижения максимума следует главным образом отнести за счет дыхания, что также наблюдали Бирекка и др. (1964), пользуясь методом меченых атомов.
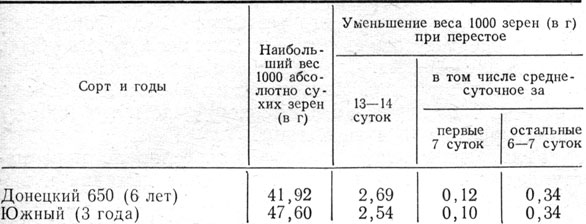
Таблица 19. Уменьшение веса 1000 зерен ячменя при перестое на корню
Естественно, что интенсивность дыхания зерна и потери органической массы на этот процесс обусловливаются степенью влажности и продолжительностью периода высокой влажности. Известно, что при влажности зерна более 14 - 15% интенсивность его дыхания и выделение углекислоты заметно возрастают, особенно при повышенной температуре. Между тем максимальный вес сухого вещества зерна ячменя достигает при его влажности около 20%. К тому же зерно, в котором незакончен период послеуборочного дозревания, дышит значительно интенсивнее, чем достигшее физиологической зрелости и находящееся в состоянии покоя (Кулешов, 1936; Прохоров и Кретович, 1951; Доброхотов, 1959). Наряду с этим зерно, созревающее на корню, в результате дождей и утренних рос медленно высыхает или повторно увлажняется; семена вбирают влагу быстро. Высушивание же зависит от относительной влажности воздуха, температуры и движения воздуха. При теплой погоде и особенно повторном увлажнении зерна резко усиливается его дыхание и увеличивается расход сухого вещества.
Установлено, что убыль веса зерна в конце созревания и при перестое происходит главным образом в результате уменьшения содержания углеводов. Анализ показал, что содержание сырого протеина в зерне ячменя возрастает до наступления твердой спелости и при перестое не уменьшается, а содержание крахмала возрастает до конца восковой спелости и затем убывает.
На ранних фазах образования зерна при дыхании выделяется много углекислоты. Например, в 1962 г-22 июня, когда влажность зерна сорта Южный на корню была 55,1%, 100 г зерна выделяли за 1 час 103,43 мг углекислоты, то есть за сутки было израсходовано 1,59 г глюкозы, что составляет 0,68 г на 1000 зерен. В этот день вес абсолютно сухого вещества 1000 зерен был 40,13 г, среднесуточный прирост составил 2,89 г, а убыль на дыхание - 0,68 г. Следовательно, есть основание считать, что в этот день приток сухого вещества составил 3,57 г на 1000 зерен, но примерно 1/5 часть его (0,68 г) была израсходована на дыхание.
Увлажнение зерна на корню, особенно по мере наступления спелости, вызывает усиление дыхания и выделение углекислоты. Так, в 1962 г. 100 г зерна сорта Донецкий 650 в середине восковой спелости (3 июля) выделили за 1 час 22,45 мг углекислоты, что составило 0,17 г сухого вещества углеводов на 1000 зерен, а при увлажнении на корню за 16 - 17 часов до отбора образцов - 43,50 мг, или 0,33 г сухого вещества. В отдельных случаях при определенных погодных условиях потери веса зерна могут быть довольно большие. Особую озабоченность вызывает тот факт, что в период интенсивного накопления веществ в зерновке значительная их часть расходуется на дыхание.
Естественно, что в период налива зерна дыхание необходимо как источник энергии для метаболических процессов, но невольно возникает вопрос, нельзя ли уменьшить расход углеводов. Если бы на ранних фазах формирования зерновки можно было уменьшить расход углеводов на дыхание, то это позволило бы значительно увеличить накопление веществ и повысить крупность, тяжеловесность зерна.
Из-за недостаточной изученности процесса зернообразования ячменя этот вопрос трактовался по аналогии с пшеницей. Однако наши исследования показали, что у ячменя наблюдаются некоторые особенности образования зерна, например трудно по темпам накопления сухой массы отделить фазу формирования зерна от налива. У пшеницы же наблюдается четкое резкое повышение притока сухого вещества после завершения формирования длины зерновки. В фазе налива у ячменя не возрастает вес сухого вещества зерна, при почти постоянном весе в нем воды. В отдельные годы прирост веса сухого вещества зерна ячменя отмечался вплоть до твердой спелости, в отношении же пшеницы этого не наблюдалось.
Как указывал Н. Н. Кулешов (1963, 1964), поступление пластических веществ в зерно прекращается при снижении его влажности до 38 - 40%. Объясняется это тем, что при такой влажности происходит коагуляция белковых коллоидов, после чего поступление сухих веществ и влаги в зерно возобновиться не может. Что же касается ячменя, то средняя влажность образца не исключает наличия части зерновок с более высокой влажностью.
Нами установлена заметная неравномерность в наступлении спелости зерен в пределах одного и того же растения и даже колоса. Например, в среднем за 3 года в конце восковой спелости (через 29 - 31 день после цветения) влажность зерна ячменя сорта Донецкий 650 была в колосе побега кущения 35,8%, а в колосе главного побега 20,9%, в том числе в верхнем ярусе 18,7% и в нижнем 23%. Подобное явление отмечается в ряде работ (Соседов и др., 1954; Доброхотов, 1959). Кроме того, увеличение веса сухого вещества зерна после наступления средней предельной влажности можно также объяснить неравномерностью обезвоживания эндосперма зерновки ячменя.
Из ряда работ следует, что эндосперм формируется и поспевает в разных местах семени неодновременно. Наиболее молодые клетки эндосперма расположены вблизи зародыша (Модилевский и др., 1958). Зародыш выделяет в 12 раз больше углекислоты, чем все семя в целом, и, следовательно, затраты углеводов на дыхание в основном сосредоточены вблизи зародыша (Леманн и Айхеле, 1936). Пополнение эндосперма, использованного зародышем в этом месте, продолжается благодаря притоку веществ из материнского организма до конца (Худяк, 1963) прекращения физиологической связи зерновки с материнским растением.
Следовательно, обезвоживание клеток эндосперма при наступлении спелости происходит не мгновенно во всей массе зерновки, а постепенно. В дорзальной (спинной) части кончика зерна эндосперм созревает раньше и его влажность может быть ниже; клетки, примыкающие к бороздке на вентральной (брюшной) стороне, ближе к зародышу, созревают последними, и их влажность может быть выше. Это подтверждается нашими данными определения влажности верхней части плода, его спинки и нижней половинки со стороны бороздки в процессе зернообразования. В связи с этим есть основание считать, что утверждение В. Доброхотова о том, что в начале восковой спелости зерно теряет связь с материнским растением, требует уточнения.
В наших опытах на Эрастовской опытной станции (1962 г.) за время от конца тестообразного состояния зерна до середины восковой спелости вес 1000 зерен ячменя сорта Донецкий 650 увеличился на 2,03 г, сорта Южный - на 1,25 г; в благоприятном 1965 г. за время от начала до конца восковой спелости вес 1000 абсолютно сухих зерен ячменя сорта Южный возрос с 46,97 до 49,66 г.
Следовательно, до конца восковой спелости вес зерновки заметно увеличивается, несмотря на то, что из-за повышенной ее влажности (более 20%) наблюдается интенсивное дыхание, на которое тратится значительное количество углеводов. Очевидно, расход сухого вещества на процессы дыхания в это время компенсируется притоком их в зерновку из вегетативных органов. В зависимости от условий, определяющих интенсивность дыхания созревающей зерновки, изменяется и прирост ее веса в этот завершающий период. Поэтому нет сомнения, что до конца восковой спелости ячменя продолжается приток веществ в зерновку. После компенсации расхода углеводов на процессы дыхания часть из них откладывается в зерновке, благодаря чему наблюдается небольшое увеличение ее веса.
При перестое ячменя вес сухого вещества зерна снижается особенно в последние дни. Имеющиеся по этому вопросу данные, а также результаты наших исследований позволяют рассматривать динамику сухого вещества в зерне как результат двух взаимно противоположных процессов: притока пластических веществ в эндосперм и частично расхода их на развитие зародыша, на дыхание зерна, а иногда и на так называемое выщелачивание и экзосмос.
Первый процесс заканчивается с исчезновением хлорофилла в вегетативных частях растений, нарушением связи между зерновкой и материнским растением и наступлением предельной влажности зерна, при которой происходит коагуляция всех содержащихся в эндосперме коллоидов. Второй процесс продолжается дольше, но интенсивность его определяется влажностью зерна, а также влажностью и температурой окружающего воздуха.
В период, когда наблюдается прирост веса зерна, очевидно, приток пластических веществ превышает его расход.
Затем постепенно наступает нулевой баланс этих двух процессов - приток равен расходу, в этот период отмечается постоянный вес зерна, что обычно называют завершением налива. В действительности же перемещение пластических веществ в зерно, пока его средняя влажность выше 15 - 16%, может еще продолжаться в размерах, равных расходу на дыхание. Без этого должна наблюдаться убыль, так как при такой влажности потери углеводов на дыхание бывают заметными. Поэтому, вероятно, одним из признаков прекращения поступления пластических веществ в зерновку является некоторая убыль веса зерна в период, когда влажность его не ниже 15 - 16% при погодных условиях, способствующих усиленному дыханию или вымыванию углеводов.
При понижении температуры, затягивании отмирания ассимилирующих органов и уменьшении расхода пластических веществ на дыхание баланс может стать положительным, что свидетельствует о продолжении поступления пластических веществ в зерновку. Этим, вероятно, и объясняется отмеченный выше факт, что в годы (или районах) с более низкой температурой в период налива и созревания, когда вегетативные органы хорошо развиты и долго сохраняют высокую влажность, прирост сухого вещества в зерне заканчивается в более поздние сроки, чем в годы (или в районах) с более высокой температурой и меньшей влажностью в этот период.
Резкое снижение веса зерновки в последние дни перестоя, по-видимому, происходит после полного физиологического отчленения зерновок, находящихся в колосе, от материнского растения. При этом обнаруживается со всей полнотой расход углеводов на дыхание зерновок, которые систематически увлажняются ночью от росы или от мелких, даже бесполезных в агрономическом отношении осадков.
В первые дни после наступления максимума веса зерна наблюдающиеся потери углеводов, вероятно, в какой-то степени еще компенсируются поступлением их из материнского растения. Если это так, то продолжительность допустимого срока уборки должна устанавливаться с учетом этого обстоятельства, так как каждый день перестоя после того, как зерно потеряло физиологическую связь с материнским растением, когда потери органического вещества резко возрастают, сопровождается ощутимыми биологическими потерями, уже не говоря о потерях от осыпания.
Всестороннее изучение процессов зернообразования с целью максимального сокращения возможных биологических потерь сухого вещества при неблагоприятных погодных условиях в период формирования и созревания имеет весьма важное значение, так как позволит уточнить приемы, обеспечивающие получение более крупного зерна.
Более высокий вес 1000 зерен при прочих равных благоприятных условиях обеспечивает больший урожай зерна ячменя. Это подтверждается данными госсортоучастков Днепропетровской области за 17 лет (табл. 20).
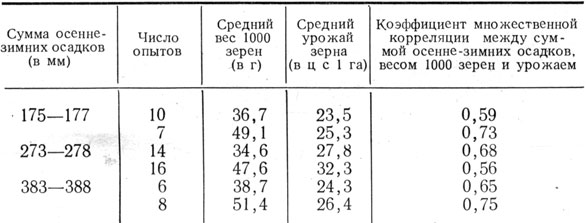
Таблица 20. Урожай ячменя в зависимости от веса 1000 зерен
Многочисленные опытные данные свидетельствуют о последовательной зависимости: чем больше вес 1000 зерен, тем выше урожай, а вес 1000 зерен тем больше, чем продолжительнее период их образования. Это опять-таки может косвенно послужить подтверждением того, что накопление массы в зерне может происходить до твердой спелости-
Ячмень в отличие от других зерновых культур обладает важной биологической особенностью создавать урожай зерна, составляющий по весу примерно половину урожая надземной массы. Отношение зерна к вегетативной массе, почти равное 1:1, характерное для большей части распространенных сортов, указывает на рациональное использование органических веществ этого растения при создании наиболее ценной части урожая - зерна.
Накопление большой вегетативной массы растениями ячменя, начиная с первых фаз роста, является необходимой предпосылкой высокого урожая зерна. Экспериментальный материал позволяет заключить, что чем лучше развиты вегетативные органы растения, тем больше образуется в его колосе зерен, чем выше урожай соломы и половы, тем, как правило, выше урожай зерна.
Однако мощно развитая вегетативная масса не во всех случаях обеспечивает высокий урожай зерна. Зерновая продуктивность растения определяется не только мощностью вегетативной массы, но и условиями, способствующими более полной мобилизации органических соединений вегетативной массы в репродуктивный период для образования зерна в колосе.
Одним из решающих условий, благоприятствующих хорошему наливу зерна, являются достаточная увлажненность и оптимальная температура в период зернообразования. Анализ данных, полученных за ряд лет, показал, что, когда в период налива зерна температура воздуха была 21 °С и выше, а относительная влажность его опускалась до 37 % и ниже, значительная часть пластических веществ оставалась в вегетативных органах и выход зерна не превышал 42%; при температуре ниже 21 °С и относительной влажности воздуха 41% и более урожай зерна в общей надземной массе достигал 53%. Так, в 1964 г., когда в период налива зерна среднесуточная температура была наиболее высокая (23,3 °С), относительная влажность воздуха в 13 часов была наименьшая (25%) и осадков не было, зерна в общей надземной массе было 33,4%, а в годы, более благоприятные (1959, 1962, 1963), - 48,7 - 53%. При неблагоприятных условиях в период зернообразования в 1964 г. значительная часть пластических веществ оставалась в вегетативных органах растений ячменя и не полностью была использована для формирования зерна.
Погодные условия в период формирования и созревания зерна трудно предвидеть. Однако важно, что отрицательного влияния мощно развитой вегетативной массы на формирование зерна не наблюдалось. При меньшей надземной массе выход зерна не возрастал. При одинаковом примерно количестве осенне-зимних осадков больший урожай зерна получался при большей вегетативной массе (табл. 21). Это подтверждается и данными Эрастовской опытной станции: при урожае общей надземной массы в 1953, 1955, 1957 гг. 43,8 - 62,5 ц с 1 га зерна в урожае было 45,5 - 48,2%, а в 1956 г. соответственно 40,5 ц с 1 га и 39,8%.
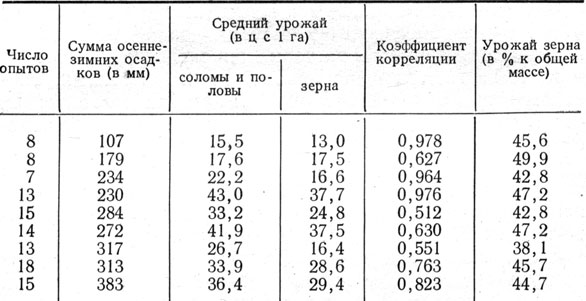
Таблица 21. Урожай зерна ячменя в зависимости от урожая надземной вегетативной массы и суммы осенне-зимних осадков по материалам госсортоучастков Днепропетровской области за 1944 - 1970 гг.
В среднем за 13 лет урожай зерна ячменя в нашем стационарном опыте лаборатории удобрений на контроле составил 17,9 ц, соломы и половы - 20,4 ц с 1 га, а на варианте с удобрением соответственно 25,9 и 31,4 ц. Зерна в урожае общей массы содержалось при удобрении 45,3%, без удобрения 46,7%.
В большинстве случаев содержание зерна в урожае общей надземной массы остается почти постоянным как при слабом, так и при сильном развитии вегетативных органов растений ячменя. При неблагоприятных условиях в период формирования и созревания зерна накопленные запасные и пластические вещества используются недостаточно как мощными, так и малоразвитыми растениями. Следовательно, способность растений ячменя наиболее полно использовать накопленные органические вещества для создания урожая зерна не нарушается и при обильно развитой вегетативной массе растений.
Обнаруживается прямая коррелятивная зависимости между урожаем зерна и урожаем вегетативной массы растений ячменя сортов Грушевский, Донецкий 650 и Южный, на что указывают величины коэффициента корреляции, вычисленного нами по данным стационарного опыта лаборатории удобрений на Эрастовской опытной станции (табл. 22).

Таблица 22. Коррелятивная связь между урожаем зерна, соломы и половы ячменя
В других агротехнических опытах (в среднем за 8 лет) коэффициент прямолинейной корреляции между урожаем надземной массы и урожаем зерна составил от 0,760+0,190 до 0,990±0,007 и в 72 опытах на госсортоучастках Днепропетровской области (18 лет) - от 0,789±0,103 до 0,910±0,065. Эти опытные данные свидетельствуют о том, что в условиях степной зоны агротехника должна быть направлена на усиление роста растений и создание с начала вегетации возможно большей надземной массы.
Можно высказать предположение, что это будет относиться и к короткостебельным сортам, так как короткостебельность не равнозначна уменьшению биомассы; у короткостебельных сортов большие резервы органического вещества откладываются в тканях стебля, благодаря чему он заметно утолщается, а также в узлах кущения и корнях.
Плод ячменя односемянный - зерновка. Крупность зерновки зависит от сорта и условий выращивания. В пределах колоса наиболее крупные зерновки формируются в средней части колоса, где цветки колосков раньше зацветают и во время налива лучше обеспечиваются питательными веществами.
У многорядного ячменя зерновки центрального колоска на каждом уступе колосового стержня симметричны и крупнее боковых несимметричных. У двухрядного ячменя зерновки всех колосков симметричны, в средней части колоса они крупнее, у основания и вершины - мельче. Поэтому в партии семенного материала двухрядного ячменя все зерновки симметричны, более выравнены и почти одинакового веса. В семенном материале многорядного ячменя приблизительно 60% зерновок несимметричных - с искривленным основанием. Изогнутые зерна меньшего размера, чем симметричные. По этим особенностям можно быстро, без сложных анализов определить, не смешаны ли в данной партии двухрядный и многорядный ячмени.
С брюшной стороны зерновки имеется продольная бороздка, в нижней части которой находится так называемая основная щетинка зерна. Эта щетинка бывает различной по степени опушения (волосистое или войлочное), величине и является одним из отличительных сортовых признаков.
При поперечном разрезе зерновки, освобожденной от цветочных пленок, различают следующие составные ее части, начиная с периферии: плодовую оболочку (перикарпий), которая образовалась из стенок завязи и состоит из двух слоев; семенную оболочку (спермодерма), развившуюся из двух оболочек семяпочки; остатки ткани ядра семяпочки (перисперм); алейроновый слой эндосперма, состоящий из нескольких рядов крупных клеток с толстыми стенками, в которых содержатся главным образом белки и жиры. Под алейроновым слоем находится внутренний эндосперм; клетки эндосперма, прилегающие к алейроновому слою, заполнены основными белковыми веществами, а остальные главным образом крахмальными зернами.
Очень важная роль в жизни семени отводится периферической зоне клеток эндосперма - алейроновому слою (Цингер, 1958). Этот слой с самого начала существования эндосперма проявляет наибольшую физиологическую активность и по содержанию важнейших питательных веществ немногим уступает зародышу ярового ячменя.
Раньше считали, что при прорастании зерна ячменя главным местом образования фермента является щиток зародыша. Однако новые исследования показали, что алейроновые клетки проявляют большую активность в синтезе фермента (альфа-амилаза), чем клетки щитка зародыша (Диксон, 1973).
В вызревшем сухом зерне ячменя содержится сухого вещества около 86 - 88% и воды 12 - 14%. Примерно 84 - 85% сухого вещества зерна приходится на органические соединения, из которых преобладают углеводы, составляющие около 80% сухих веществ. Основной углевод в зерне ячменя - крахмал,
По данным Государственной комиссии по сортоиспытанию сельскохозяйственных культур при Министерстве сельского хозяйства СССР, в зависимости от сорта, года урожая и места произрастания содержание в зерне крахмала колеблется в довольно широких пределах - от 48 до 67%, а белка - от 8 до 22%.
Оболочки зерновки, выполняя функции покрова, одновременно являются важной ее составной частью, играющей существенную роль в формировании зерна и жизнедеятельности зародыша. В наших опытах на Эрастовской опытной станции в 1964 г. в течение семи дней до тестообразного состояния зерна ячменя сорта Южный вес эндосперма десяти зерновок средней части колоса увеличился на 0,245 г, зародыша на 0,014 г, а цветочной и плодовой оболочек уменьшился на 0,099 г. Очевидно, такое изменение веса оболочек связано с выявленной (Риявен, Бранбури, 1960) доминирующей ролью семенной оболочки в синтезе глутамина и распределении других энзимов в зерновке. Вероятно, этим объясняется и наблюдаемое А. М. Мироненко (1953) уменьшение веса зерна ячменя в результате повреждения покровов цветка при скрещивании.
Ф. М. Куперман (1964) считает, что в призародышевой части спинки семени локализуются физиологически активные ростовые вещества, регулирующие силу роста и направление зародышевых листочков. Таким образом, предохранение покровов семени от повреждений имеет большое значение в сохранении его семенных достоинств. Велика роль оболочек во время хранения семян и их прорастания. Поэтому при уборке и очистке нельзя допускать повреждения оболочек зерна. На спинной стороне зерновки у ее основания расположен зародыш. Он состоит из почки (зародышевые листья и конус нарастания), прикрытой колеоптиле, и зародышевых корешков, защищенных колеоризой. Эти две главные части зародыша отделены зародышевым стебельком, соединяющимся со щитком. Щиток примыкает к эндосперму и выполняет роль органа, всасывающего питательные вещества из эндосперма и направляющего их в зачатки растущих органов зародыша, В эндосперме зерновки содержатся все питательные вещества для начального развития растения до перехода его на самостоятельное питание.
Семенные качества зерновки определяются также условиями зернообразования. Гидротермические условия формирования и созревания семян оказывают большое влияние на их посевные и урожайные свойства. Повышенная температура ускоряет, а пониженная задерживает процесс развития и дифференциации зародыша семени (Оксиюк и Худяк, 1958; Абрамова, 1961). Известно (Валишина и Цингер, 1952), что чем лучше развиты органы зародыша, тем выше его жизнеспособность, тем скорее зерновка, при соответствующих условиях, может прорастать и обеспечивать в дальнейшем образование полноценного растения.
В наших опытах по мере созревания возрастал вес зерновок, параллельно с этим повышались их полевая всхожесть и вес надземной массы растений (табл. 23).
Из данных таблицы можно заключить, что дозревание семян, особенно на ранних фазах уборки, заключается не только в приобретении способности прорастать, то есть в преодолении периода покоя, но, вероятно, и в дальнейшей дифференциации зародыша. Иначе чем же объяснить, что при использовании дозревших зерновок вес сухой массы растений, достигших фазы кущения, резко возрастает. Очевидно, при дозревании зерновок, особенно на материнском растении, они еще некоторое время получают дополнительное питание и продолжается приток веществ гормонального характера из свежего колоса, благодаря чему дифференциация и развитие зародыша протекают дольше, чем при немедленном после уборки отчленении.
Наклевывание и образование главного корешка в первые часы прорастания при дозревании зерен в колосе на материнском растении при раздельной уборке протекали более интенсивно, чем при прямом комбайнировании.
Улучшение посевных качеств семян, убранных в ранние фазы спелости и дозревших в колосе или в снопах, отмечается во многих работах. Процессы, обусловливающие улучшение посевных качеств, полнее протекают при медленном высыхании хлебной массы, так как удлиняется период жизнедеятельности семян на скошенных растениях. При этом большое значение имеют степень оводненности вегетативной массы, влажность стеблей при скашивании и гидротермические условия, складывающиеся в валках.
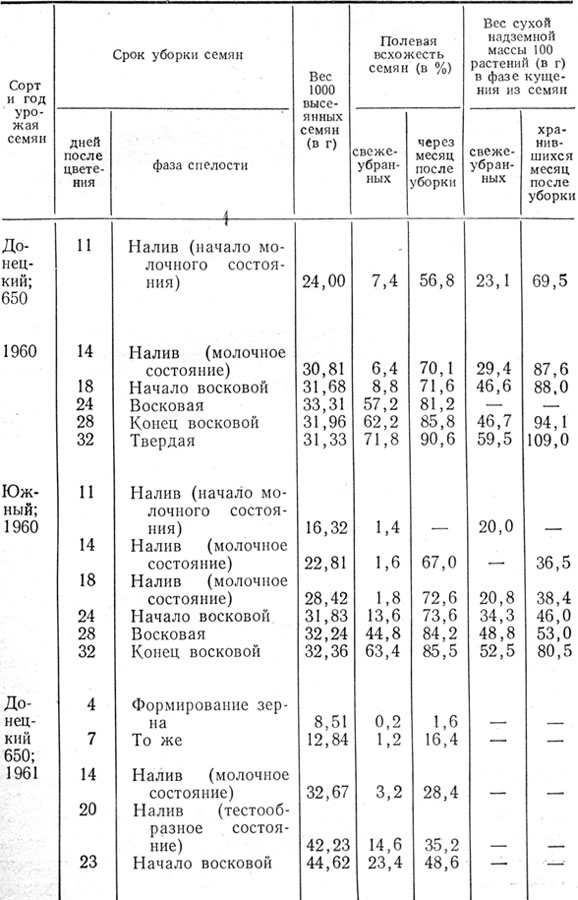
Таблица 23. Полевая всхожесть и вес надземной массы растений из свежеубранных и дозревавших разновозрастных семян ячменя
Данные специального эксперимента показали, что искусственное увлажнение (30 мм) валков вслед за скашиванием с целью изменения гидротермического режима приводило к некоторому повышению относительной влажности и снижению температуры воздуха в скошенной массе: через двое суток после скашивания относительная влажность воздуха вечером была в обычном валке 70%, в увлажненном - 97%, а температура соответственно 21,2 и 20,3 °С тепла. Это, естественно, несколько замедлило высыхание вегетативной массы и зерна, продлило срок их физиологической связи, в результате чего зародыш дольше развивался, чем в обычных валках.
Дозревание растений в искусственно увлажненных валках после скашивания в ранние фазы, как правило, не приводило к снижению полевой всхожести, а, наоборот, в отдельные годы даже несколько ее повышало. Например, в 1958 г. полевая всхожесть семян урожая 1957 г., убранных в начале и середине восковой спелости и дозревавших в обычных валках, составляла 76,6 - 78%, а дозревавших в увлажненных валках - 84,5 - 86,6%. При этом всходы из семян, дозревавших в увлажненных валках, появились более дружно; в первые дни подсчета их было на 6,4 - 6,5% больше, чем из семян, дозревавших в обычных валках.
При дозревании в увлажненных валках растения из таких семян меньше поражались пыльной головней, чем при дозревании в обычных валках. В 1960 г. в посеве из семян, убранных в 1958 г. в середине восковой спелости и дозревавших в обычных валках, оказалось 3,4% растений, пораженных пыльной головней, в посеве из семян, дозревавших в увлажненных валках,- 2,1%. Пораженных пыльной головней растений в среднем за 4 года было соответственно 2,2 и 1,2%.
Замедленное высыхание скошенных растений в увлажненных валках, по-видимому, способствует лучшему дозреванию семян и более полному развитию зародыша. Очевидно, из семян с полнее развитым зародышем формируется растительный организм, обладающий повышенной жизненностью и отличающийся рядом преимуществ, в том числе и физиологическим свойством лизировать мицелий гриба пыльной головни. Хотя различна в степени поражения небольшие, но все же наблюдается четкая тенденция к уменьшению числа пораженных растений при использовании семян, дозревавших в увлажненных валках. Влияние повышенной жизненности семян на снижение пораженности растений ячменя пыльной головней отмечал Е. Л. Куцевол (1957) и гельминтоспориозом - А. И. Ермоленко (1946).
Из вышеизложенного можно допустить, что опасение скашивать на семенных участках растения, еще не совсем высохшие на корню после дождя, не всегда оправдано. В отдельных случаях такое скашивание может оказаться более полезным, чем выжидание полного просыхания хлебостоя, однако это требует специального и более глубокого изучения.
Растения из семян, дозревавших в валках, быстрее кустятся. В 1961 г. вес 1000 семян сорта Донецкий 650 при уборке в фазе восковой спелости в день скашивания был 32,1 г, после высыхания в валках - 32,5 г, то есть практически одинаковый. Но при высеве в 1962 г. на 23 апреля растений из семян, немедленно обмолоченных, раскустилось 95,7%, а из семян, дозревавших в валках,- 100%.
При уборке в конце восковой спелости сорта Донецкой 650 при прямом комбайнировании и раздельным способом вес 1000 зерен был почти одинаковый - 45,5 и 45,8 г, но при первом подсчете взошло растений соответственно 27,5 и 38,4%, при втором - 66,1 и 79,9%, при третьем - 73,5 и 83,1% к числу высеянных семян. Из семян, убранных раздельным способом, растения не только быстрее начинают куститься, но и раньше и полнее выколашиваются.
Семена ячменя сорта Южный, убранные в 1964 г. раздельным способом в конце восковой спелости, дали в 1965 г. более урожайные растения, чем убранные прямым комбайнированием при твердой спелости (табл. 24).
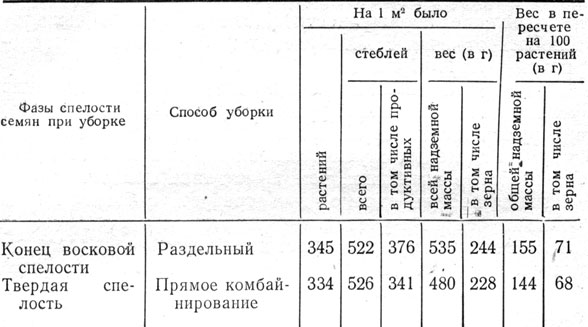
Таблица 24. Урожайные качества семян ячменя сорта Южный в зависимости от срока и способа их уборки
Аналогичные данные получили также Л. К. Мурадханян, Н. Н. Капитаненко (1957), И. Г. Строна (1962), С. Ф. Архангельский, В. В. Шелепов (1965).
Различные погодные условия в период зернообразования, помимо влияния на накопление сухих веществ и, следовательно, на образование эндосперма, по-разному сказываются на развитии зародыша, содержании азота и таким образом на посевных и урожайных качествах семян. Благоприятные условия корневого питания способствуют открытому цветению (Секун, 1963; Довбах, 1964), а при открытом цветении семена отличаются более высокой энергией прорастания и образуют более продуктивное растение, чем при закрытом цветении (Тетерятченко, 1964).
При опылении в оптимальный срок формируются семена, из которых образуются более продуктивные растения, чем при опылении слишком молодой или стареющей яйцеклетки (Коренев, 1959; Львова, 1960; Удольская, 1963). Глубокое воздействие на посевные и урожайные качества семян оказывает место их формирования в колосе, так как цветение, оплодотворение, налив и дальнейшее развитие зародыша зерновки в разных ярусах колоса протекают неодновременно.
В наших опытах выявлено, что во время молочного состояния у сорта Донецкий 650 вес сырого вещества десяти зерновок среднего яруса составил 0,768 г, в том числе эндосперма 0,537 и зародышей 0,020 г, а нижнего и верхнего ярусов соответственно 0,684 - 0,582; 0,419 - 0,393 и 0,016 - 0,017 г. У сорта Южный вес зерновок среднего яруса был 0,870 г, в том числе эндосперма 0,562 и зародышей 0,025 г, а нижнего и верхнего ярусов соответственно 0,786 - 0,596; 0,486 - 0,400 и 0,018 - 0,012 г. Это согласуется с данными и других исследователей (Куперман, 1950; Паршакова, 1964; Коновалов, Случевская, 1965; Фишбекк, 1965), свидетельствующих, что в центральной части колоса семена начинают формироваться раньше, накопление в них веществ протекает энергичнее, они наиболее полновесные и отличаются большим весом зародыша. Такие семена дают более мощные проростки (Коновалов, Удалов, 1964), и растения из них меньше поражаются пыльной головней (Демирликакмак и др., 1963). Растения из семян, сформированных в средней части колоса, отличаются более высокой продуктивностью (Мухин, 1941; Колганов, 1949; Абхазова, 1950; Максимчук, 1951; Строна, 1964).
Фазы роста и развития. Среди яровых колосовых ячмень наиболее скороспелая культура. Как растение длинного дня, ячмень для своего развития требует сравнительно длительного освещения. Поэтому в северных районах период от всходов до колошения меньший, чем на юге, где световой день короче. Продолжительность вегетационного периода зависит от биологических особенностей сорта и места его выращивания. Длина вегетационного периода ячменя в условиях СССР, по данным государственного сортоиспытания за 10 лет, колеблется от 72 дней в Якутской АССР до 121 дня в Дагестанской АССР.
Различают следующие фазы развития растений ячменя: появление всходов, кущение, выход в трубку, колошение и спелость. В наших опытах на Эрастовской опытной станции в среднем за 15 лет продолжительность периода от посева до всходов была 17 дней, от всходов до кущения - 22, от кущения до колошения - 24, от колошения до молочного состояния зерна - 15 и от молочного состояния до твердой спелости - 14 дней. Таким образом, в условиях Днепропетровской области период вегетации ячменя (от всходов до спелости) в среднем составляет 75 дней.
В жизни растения весьма ответственным является период образования генеративных органов. Колошение ячменя начинается с выхода остей из пазухи верхнего листа и частично колоса из боковой щели листа. При пасмурной погоде во время колошения колосовой стержень растягивается и колос вырастает рыхлый, а при интенсивном солнечном освещении колос формируется более плотный. Ячмень - растение самоопыляющееся. Пыльники и рыльца в колосках созревают одновременно еще до выхода колоса из пазухи листа, и потому пыльца попадает на рыльца своего же цветка. Правда, в условиях засухи бывает и открытое цветение ячменя, и, следовательно, возможно также перекрестное опыление.
Процесс зернообразования делится на три периода: формирование, налив и созревание, в свою очередь, включающие соответствующие фазы (Кулешов, 1963, Строна, 1966).
По девятилетним данным наших исследований, у ячменя период формирования продолжается 10 - 11 дней от оплодотворения до начала молочного состояния зерновки и включает фазы роста и водянистого ее состояния. В это время увеличивается завязь, зерновка в основном достигает окончательной длины и содержит водянистую жидкость. К концу периода зерновка отличается зеленой окраской, содержит 36 - 39% сухого вещества (от максимального веса) и 68 - 70% воды; в это время засыхают пластинки нижних листьев.
Период налива продолжается примерно 11 - 14 дней от начала молочного до конца тестообразного состояния зерна и включает фазы предмолочную, молочную и тестообразную. В этот период происходит наибольшее накопление минеральных и органических веществ; к концу периода консистенция эндосперма становится тестообразной, вес сухой массы зерновки достигает наибольшей его величины - 93 - 95%, влажность 46 - 48%. Зародьшг увеличивается и становится хорошо заметным, значительно увеличиваются ширина и толщина зерновки, окраска ее в основном светлеет; зеленая окраска остается лишь возле зародыша и в бороздке. Влагалища листьев нижних ярусов желтеют, зелеными остаются узлы стебля, листья верхнего яруса и колосья.
Период созревания начинается после окончания тестообразного состояния зерна и включает фазы восковой и твердой, или, как часто выражаются, уборочной, спелости. Фаза восковой спелости ячменя продолжается 5 - 8 дней и имеет весьма важное производственное значение. Для правильного определения оптимальных сроков уборки раздельным способом следует различать начало, середину и конец восковой спелости.
В начале восковой спелости зерно крупное, поверхность его блестящая, желтого цвета и полностью лишена зеленой окраски, легко режется ногтем. При надавливании содержимое зерновки не извлекается, мнется, как воск, но к пальцам не пристает. Основная масса растений желтая, лишь у части растений верхние стеолевые узлы и колосья зеленые; влажность зерна 35 - 40%.
В середине восковой спелости зерновка ногтем еще режется, эндосперм белый, мучнистый или стекловидный, между пальцами не скатывается. Благодаря обезвоживанию размер зерновки в сравнении с началом фазы уменьшается, влажность ее 25 - 35%. Растения полностью желтеют, но стебли эластичные, неломкие.
В конце фазы восковой спелости зерновка ногтем не режется, от надавливания остается лишь вмятина - слабый след ногтя. Влажность зерна 21 - 22%, размер и окраска его такие же, как и твердой спелости данного сорта. Зерно из колосков не выпадает.
Продолжительность фазы восковой спелости определяется агротехническими и погодными условиями. В наших опытах на Эрастовской опытной станции в засушливом 1957 г. восковая спелость ячменя сорта Донецкий 650 продолжалась 5 дней, в 1958 г.-7; в засушливом 1962 г. восковая спелость сорта Южный продолжалась 4 дня, в 1965 г. - 9 дней.
В литературе распространено мнение о том, что к началу восковой спелости заканчивается приток веществ в зерновку. В отношении ячменя ярового это экспериментально не подтверждается. Результаты исследований в условиях степной зоны Украины позволяют заключить, что в фазе восковой спелости физиологическая связь зерновки с материнским растением и приток веществ в зерновку продолжаются. Следовательно, при слишком ранней уборке преждевременно останавливается поступление веществ в зерновку. Уборка прямым комбайнированием до конца восковой спелости, когда процессы дыхания зерновки протекают интенсивно, приводит к значительному недобору урожая.
Твердая спелость продолжается 2 - 3 дня. В начале твердой спелости зерновка становится твердой, ее можно только ломать или разрезать, и приобретает размер, форму и окраску, характерные для сорта. Если в зерне содержится 17 - 22% влаги, можно убирать прямым комбайнированием, но до хранения его требуется просушить. В конце твердой спелости стебли приобретают соломисто-желтый цвет, засыхают, становятся ломкими, зерно может осыпаться. При перестое на корню урожай снижается как из-за осыпания, так и в результате биологических потерь, вызывающих снижение веса зерна, остающегося в колосьях. Полная спелость зерна наступает после прохождения периода послеуборочного дозревания.
Отношение ячменя к условиям произрастания. Высокие урожаи ячменя обеспечиваются при создании среды, наиболее полно отвечающей требованиям растений на протяжении всего периода вегетации. Требования растений к условиям существования определенным образом отражают условия, в которых они возникли.
Развивая учение Н. И. Вавилова о возникновении и введении в культуру ячменя, А. Я. Трофимовская (1971) на основании новейших данных уточнила и охарактеризовала центры происхождения ячменя (генцентры) в связи с локализацией в них важнейших селекционных признаков его ботанических разновидностей. А. Я. Трофимовская считает, что весьма широкий ареал распространения ячменя обусловлен огромным полиморфизмом культуры и исключительным богатством его экологических типов.
Например, на обширной территории европейско-сибирского генцентра произошла дифференциация экотипов по реакции на условия среды - от типичных гидрофилов Западной Европы до засухоустойчивых степных рас юга Украины, Северного Кавказа, Юго-Востока Российской Федерации, Западной и Восточной Сибири. Следовательно, на территории, где теперь размещается около половины посевов ячменя в нашей стране, эволюция рода гордеум совершалась в направлении эфемеризации видов культуры ячменя. Так, степная и лесостепная экологические группы сортов ячменя ярового являются среднеспелыми и среднепоздними. Первые отличаются повышенной засухоустойчивостью на протяжении всего вегетационного периода и на коротком дне задерживают колошение в средней степени, вторые - средней засухоустойчивостью на ранних фазах и на коротком дне задерживают колошение в сильной степени (Трофимовская, Бахтеев, 1966).
Динамичность в период вегетации и периодические закономерные изменения почвенно-климатических условий в процессе эволюции ячменя обусловили формирование наблюдаемых теперь: биологических и физиологических особенностей этой культуры. Н. И. Вавилов (1957) считал, что физиологические и биологические особенности растительного организма тесно связаны с условиями среды.
Эфемеризация ячменя в условиях ксерофилизации климата и возникновение засухоустойчивых степных рас, отличающихся способностью раньше созревать и в засушливых условиях создавать урожай в основном за счет осадков, накопленных в почве к началу вегетации, существенным образом сказались также на характере корневого питания растений.
Под влиянием среды в процессе эволюции сложились определенные требования растений ячменя не только к условиям увлажнения и продолжительности светового дня, но и к режиму питания и аэрации (Разумов, 1950). Например, растения ячменя степного экологического типа приобрели способность на ранних фазах развития запасаться элементами минеральной пищи и в дальнейшем за их счет формировать нормальный урожай.
Периодичность в минеральном питании растений в определенной мере как бы отражает наличие легкодоступных их форм в почвенном растворе. Это легко проследить на примере динамики фосфора в черноземных почвах засушливых и полузасушливых районов. Например, в ряде работ (Щусьев, 1902; Кирсанов, 1937; Францесон, 1949; Вернандер и др., 1951; Попов, Дмитренко, 1951; Лебедянцев, 1960) выявлено, что на черноземах содержание общих запасов фосфора и легкоподвижных его форм по мере углубления по почвенному профилю уменьшается.
По результатам исследований С. М. Драчева (1928), Ф. В. Трифонова (1929), А. В. Соколова и Т. Д. Корицкой (1948), С. Тандилль (1959), Э. И. Шконде (1960), М. Стратулат (1972) есть основание заключить, что весной благодаря повышенной влажности почвы фосфатов в почвенном растворе содержится больше, а с наступлением лета абсолютное количество их заметно уменьшается.
Наряду с этим установлена (Туева, 1929; Бренчли, 1929; Стрельникова, 1937; Авдонин, 1939) способность растений ячменя запасаться необходимым для них количеством фосфора в первой половине вегетации.
При сопоставлении нетрудно заметить, что содержание абсолютного количества легкодоступных фосфатов в почвенном растворе во времени (в течение вегетационного периода) и в пространстве (по почвенному профилю) изменяется с потребностью в них растений ячменя в процессе роста. Это относится и к другим элементам питания. Подтверждается это указанием А. Ф. Тюлина (1957) на то, что изменение гидротермических условий в почве в течение вегетационного периода влечет за собой качественные изменения в почвенной структуре, что, в свою очередь, отражается на изменении почвенного плодородия, создании определенных ритмов. Растения же в своем росте приспособляются к ритмам жизни почвы, создавая соответственно свои ритмы поглощения питательных веществ из почвы.
Таким образом, периодичность в минеральном питании растений следует рассматривать как отражение в структуре растительного организма ячменя той среды, в которой он развивался. Учет этих особенностей может иметь важное значение при решении практических задач растениеводства. Например, повышенные требования в начале вегетации к минеральному питанию, возникшие в процессе эволюции у растений ячменя, легко удовлетворить путем локального припосевного внесения удобрений, что, как показала практика, обеспечивает заметное повышение их продуктивности.
Требования к почве. Сжатые сроки поглощения элементов минеральной пищи в начальные фазы развития и относительно слабая усвояющая способность корней ячменя обусловливают высокую требовательность его к почвенному плодородию. Наиболее пригодны для ячменя в нечерноземной зоне суглинистые почвы средней связности, в южных районах страны черноземы. Непригодны для ячменя кислые, торфянистые почвы. Такие почвы могут быть использованы под посев ячменя лишь после известкования. Правда, в настоящее время в Белорусской ССР районирован и широко распространен сорт ячменя Полесский, который лучше других переносит повышенную кислотность, переувлажнение почвы, устойчив к полеганию и хорошо удается на осушенных торфяноболотных почвах.
Плохо растет ячмень на песчаных и супесчаных почвах. Такие почвы обычно мало содержат усвояемых питательных веществ. Лишь при внесении достаточного количества органических и минеральных удобрений такие почвы можно при необходимости отводить под посев ячменя. Совсем непригодны для ячменя легкие маловлагоемкие почвы.
Важнейшим условием получения высоких урожаев ячменя является обеспечение молодых растений достаточным количеством легкодоступной пищи, что достигается размещением его посевов на более пригодных для него почвах по лучшим предшественникам и применением удобрений.
Требования к влаге. Семена ячменя начинают прорастать при влажности, равной двойной гигроскопической влагоемкости данной почвы. Чем быстрее и больше воды семена поглощают при набухании, тем скорее они прорастают, тем выше у них энергия прорастания и сила роста (Левицкий и др., 1963).
Поглощенная вода медленно распространяется к внутренним слоям семени, она задерживается в верхних слоях, где обладает большой подвижностью и может легко обмениваться (Лебедев, 1959). Поэтому процесс поступления воды в семя не является односторонним.
Во время набухания и в течение всего периода прорастания семена ячменя поглощают отдельные вещества из окружающего их раствора (аммиак, амиды) и одновременно выделяют в раствор избыточные продукты. Выщелачиваются преимущественно растворимые углеводы (сахар, декстрин и др.), которые представляют благоприятную среду для развития плесневых грибов (Алов, 1946).
В. Н. Доброхотов (1957) показал, что при влажности ниже уровня, необходимого для прорастания, в семенах протекает процесс гидролиза белков. В результате этого накапливаются промежуточные соединения и аммиак, вызывающие нарушение функциональных процессов в клетках семени. Это тормозит деление клеток, что отрицательно сказывается на прорастании семян, силе начального роста и полноте входов. Поэтому перерывы с прорастанием семян в почве, а следовательно, длительная задержка с появлением всходов сопровождаются гибелью части зерен. На черноземных почвах запасы продуктивной влаги в количестве 15 мм в пахотном слое обеспечивают своевременное появление и удовлетворительное состояние всходов (Вериго и Разумова, 1963).
Среди хлебных злаков ячмень считается наиболее засухоустойчивой культурой. Вместе с тем он весьма требователен к увлажнению почвы в период стеблеобразования и колошения. Недостаточная влагообеспеченность, высокая температура и сухость воздуха в начале вегетации ведут к снижению густоты всходов, слабому укоренению растений, закладке небольшого количества члеников зачаточного колоса, в результате чего образуется колос с малым количеством колосков. Недостаток влаги в период образования репродуктивных органов оказывает губительное действие на пыльцу ячменя. Стерильность части пыльцы обусловливает увеличение числа бесплодных колосков и снижение продуктивности растений.
В засушливых районах особо важную роль для ячменя играют достаточные запасы влаги, накопленные в почве к началу его вегетации. Опытами Эрастовской опытной станции выявлено, что из общего количества влаги, израсходованной ячменем за вегетационный период из весенних почвенных запасов, при размещении его после кукурузы (среднее за 11 лет) использовано 77,2%, после озими (среднее за 6 лет) - 68,5%. Как правило, в годы с малым количеством осадков в период вегетации ячменя в общем расходе влаги удельный вес использованной из почвенных запасов заметно возрастает. Так, в 1961 г., когда за время вегетации ячменя осадков было 199,7 мм, из весенних запасов использован 51% влаги, а в 1957 г., когда их было 60,4 мм, расход влаги из весенних запасов составил 89,7%. В данном случае урожай ячменя (50,2 ц общей массы с 1 га, в том числе 23,1 ц зерна) получен в основном за счет запасов влаги, накопленных в почве к моменту посева.
Определение потребности ячменя во влаге по общему ее расходу в какой-то степени условно, так как в действительности взаимоотношение растений с окружающей средой весьма сложное. Например, установлено (Новицкая, 1960; Коровина, Новицкая, 1962), что при пониженной температуре растения ячменя снижают расход воды на создание единицы сухого вещества.
Значение влаги в каждый отдельный период развития определяется не только общим количеством потребления. Известно, например, что общее потребление воды на начальных фазах развития невелико. Вместе с тем недостаточное увлажнение в это время приводило к уменьшению количества зерен в колосе (Лерман, 1963). По данным В. А. Равич (1936) и И. И. Белякова (1964), особое значение для ячменя имеет достаточное увлажнение в период выход в трубку - колошение.
В наших опытах на Эрастовской опытной станции установлено, что если в период от посева до выхода в трубку ячменем используется 20 - 29% и от колошения до уборки - 12 - 25% общего расхода влаги из почвы, то от выхода в трубку до колошения потребляется 32 - 37%. В период от выхода в трубку до колошения ячмень в наших опытах поглощал влагу преимущественно с глубины 20 - 80 см, а после колошения - 80 - 120 см.
Весьма важная роль принадлежит влаге в пахотном слое во время образования узловых корней. Образование хорошо развитых вторичных корней в последующем обусловливает возможность полноценного использования почвенного плодородия, в том числе и влаги для развития более продуктивных растений.
В засушливых и полузасушливых районах страны главным источником водоснабжения ячменя в течение всего периода вегетации является влага, накопленная в корнеобитаемом слое к моменту посева. Данные 75 опытов, проведенных в течение 17 лет на государственных сортоучастках Днепропетровской области, показали, что по мере увеличения суммы осенне-зимних осадков урожаи зерна ячменя неуклонно повышаются. Когда за период с августа предыдущего года по- март выпало осадков до 100 мм, средний урожай ячменя составил 8,8 ц с 1 га, а при 261 - 300 мм - 30,2 ц. Коэффициент корреляции между количеством осенне-зимних осадков и урожаем зерна ячменя равен 0,950+0,001. Наличие высокой коррелятивной зависимости между влажностью почвы при посеве и урожайностью ячменя установил также Харрис (1963) в условиях штата Канзас в США.
Из данных госсортоучастков Днепропетровской области за 17 лет видно (табл. 26), что в годы, когда за осенне-зимний период выпадает мало осадков (не более 300 мм), урожай зерна ячменя получен тем выше, чем больше их было в период вегетации. При обильном увлажнении почвы к началу вегетации увеличение осадков вегетационного периода мало сказывается на повышении урожая.
В степной зоне при учете складывающихся погодных условий и биологических особенностей ячменя можно уже во время сева в какой-то мере предвидеть, насколько благоприятными будут условия развития культуры в текущем году.
При накоплении максимальных запасов влаги в почве ко времени посева ячменя и правильной агротехнике посева создается возможность получать хорошо развитые, жизнеспособные, с высокой продуктивностью растения, обеспечивающие неплохие урожаи даже в годы с относительно засушливым летом.
Таблица 25. Роль осадков вегетационного периода в повышении урожая ячменя в зависимости от суммы осенне-зимних осадков
Наряду с этим ввиду неравномерного выпадения осадков в течение года для получения высокого урожая фуражного зерна целесообразно регулировать набор культур с такими биологическими особенностями, которые более эффективно позволяют использовать влагу, поступающую от осадков, в течение всего периода вегетации. Если поздние культуры хорошо используют осадки второй половины вегетационного периода, то ячмень максимально использует для формирования урожая осадки осенне-зимнего, ранневесеннего и первой половины вегетационного периода.
В результате больших отклонений погодных условий от среднемноголетних показателей, даже при полном соблюдении агротехники, урожай ячменя в сравнении с другими полевыми культурами часто меняется. Например, по данным государственных сортоиспытательных участков Днепропетровской области в среднем за 14 лет, урожай кукурузы составил 30,1 ц, а ячменя - на 4,7 ц с 1 га меньше. Однако преимущество кукурузы перед ячменем наблюдается не во все годы. Так, в 1958 г. средний урожай зерна кукурузы на упомянутых сортоучастках получен 48,6 ц, выше среднего урожая зерна ячменя на 14,6 ц, а в 1939 г. средний урожай ячменя составил 32,6 ц, выше кукурузы на 16,5 ц.
В среднем за 16 лет (1958 - 1973) на Новоодесском госсортоучастке Николаевской области средний урожай зерна ячменя составил 27,4 ц с 1 га, такой же, как и кукурузы; в среднем, за 8 лет урожай ячменя был равен урожаю кукурузы или немного выше его. На Синельниковской селекционно-опытной станции из 27 лет урожай зерна ячменя был выше или равен урожаю кукурузы в 13 лет.
Правильное сочетание посевов этих культур в соответствующих зонах следует рассматривать как важнейшее средство более полного использования атмосферных осадков для выращивания фуражного зерна.
Требования к теплу. Ячмень отличается небольшой требовательностью к температуре; отдельные сорта мирятся с малым количеством тепла за вегетационный период, что обусловливает широкое распространение культуры до северных границ земледелия и довольно высоко в горах, до 4000 м над уровнем моря. Семена ячменя могут прорастать при низкой температуре 1 - 2°С тепла, что позволяет высевать их в ранние сроки.
Однако при этом прорастание сильно растягивается. Жизнеспособные всходы можно получить лишь при температуре не ниже 4 - 5°С тепла, но появление их при этом задерживается. Для образования всходов ячменя сумма активных температур (со среднесуточными выше минимальных для образования всходов) составляет 100 °С.
При более высоких температурах поглощение воды семенами происходит с большей энергией и более быстрыми темпами, чем при пониженных. Лефевр (1959) установил, что повышение температуры от 6 до 25 °С приводит к усилению всасывания воды, дыхания семян ячменя и потере сухого вещества. При пониженной температуре семена для прорастания поглощают меньше воды (Скрипчинский и Руденко, 1949). Обусловлено это также и тем, что, как установлено А. Н. Соколовским (1956), при понижении температуры возрастает водоудерживающая способность почвы. Поэтому при низкой температуре недостаток почвенной влаги особенно сильно задерживает появление всходов. При быстром нарастании температур весной, что часто наблюдается в южных районах страны, ухудшаются условия кущения и формирования зачаточного колоса.
При невысокой положительной температуре (ранней весной) период посев - всходы бывает более продолжительным. При пониженной температуре в ночное время наблюдается интенсивный рост растений, что способствует формированию растений с высокой продуктивностью и получению более высокого урожая. Это подтверждается 13-летними данными госсортоучастков Днепропетровской области (табл. 26).
Таблица 26. Зависимость урожая зерна ячменя от среднесуточной температуры воздуха в период посев - всходы
В пределах каждой группы опытов по сумме осенне-зимних осадков большие урожаи зерна ячменя получены при меньшей среднесуточной температуре периода посев - всходы, то есть при более раннем сроке сева и как следствие повышенной продолжительности периода. Поэтому крайне важно предпосевную обработку почвы и посев проводить в ранние и максимально сжатые сроки, чтобы полнее использовать весенние запасы влаги пахотного слоя и создать условия для прохождения первых этапов роста до наступления высоких температур.
В период посев - всходы часто наблюдаются утренние заморозки. Однако кратковременное похолодание существенным образом не влияет на изменение температуры почвы на глубине заделки семян. Но и при некотором снижении температуры на непродолжительный срок это не может сказаться отрицательно на полевой всхожести. Более того, исследования Е. Я. Суманова и др. (1962) показали, что небольшое промораживание (-4°С) семян ячменя в течение трех суток приводило к повышению энергии прорастания и всхожести.
Всходы ячменя безболезненно переносят заморозки 7 - 10 °С, а позже, в фазу цветения растения, повреждаются и частично гибнут при температуре минус 1 - 2°С, в фазу молочного состояния зерна -2 - 4°С (Разумова, 1968). Минусовые температуры отрицательно влияют на рост вегетативной массы в первые дни после появления всходов. На Эрастовской опытной станции в 1940 г. всходы ячменя появились 7 мая, а 12 - 17 мая были утренние заморозки 5 - 6°С. В результате этого кончики первых листьев ячменя пожелтели, затем эти листья полностью погибли, однако растения в дальнейшем нормально развивались. В 1948 г. на Магдалиновском госсортоучастке в третьей декаде апреля - после появления всходов заморозки доходили до 9,4 °С. Хотя листочки растений несколько и пострадали, все же урожай зерна в этом году получен 23,1 ц с 1 га. В 1949 г. после появления всходов ячменя сорта Донецкий 650 на Синельниковском госсортоучастке были заморозки до 4,2 °С, а урожай зерна составил 29,7 ц с 1 га.
Интересно, что на Эрастовской опытной станции на фоне рядкового удобрения растения от снижения температуры до 5,6 °С совсем не пострадали. Отсутствие повреждений растений ячменя от заморозков до 4° С отмечает Бояновски (1963) в опытах, проведенных в Польше. Обусловлено это тем, что во время заморозков конус нарастания находится не на поверхности почвы, а на глубине залегания семян, где температура почти не снижается. На дневной поверхности находятся первые листья, функции которых после гибели выполняют вновь появляющиеся. Такие повреждения утренними заморозками не сказываются отрицательно на дальнейшем развитии растений и их продуктивности, если условия произрастания благоприятные.
Опасной является высокая температура в сочетании с низкой относительной влажностью воздуха и сильными ветрами. Такое сочетание элементов погоды - суховей в южных районах страны чаще всего наблюдается в начальный период вегетации и в конце созревания ячменя. Суховеи в апреле и мае приводят к быстрому высыханию верхнего слоя почвы, что отрицательно сказывается на полноте всходов, затем на формирований узловых корней растений и урожае. По многолетним данным госсортоучастков Днепропетровской области, когда в период всходы - кущение среднесуточная температура воздуха была 11,4° С тепла (15 опытов), урожай ячменя составил 26,8 ц с 1 га, а при температуре 15°С (11 опытов) узловые корни развивались слабо и урожай был 13,1 ц.
Как установил А. М. Шульгин (1940), важнейшим фактором кущения хлебов при хорошем увлажнении является температура почвы на глубине залегания узла кущения. В 1966 г. в ряде районов степи Украины в период кущения ячменя было резкое повышение температуры. По данным Синельниковской метеостанции, в первой декаде апреля среднесуточная температура воздуха в сравнении со среднемноголетней была выше на 4,3 °С и во второй декаде на 4,9 °С тепла. По данным Комиссаровской метеостанции, относительная влажность воздуха 26 апреля была 23%, 4 мая - 20, 5 мая - 18 и 9 мая - 11%. В таких условиях в верхнем слое почвы быстро повысилась температура.
Образование узловых корней тормозилось повышенной температурой воздуха и сухостью верхнего слоя почвы. Однако и в этих условиях на отдельных участках, где вносили удобрения, узловые корни развивались энергичнее и быстрее проникали вглубь до высыхания верхнего слоя. Обусловлено это, как выявлено нашими опытами на Эрастовской опытной станции, тем, что при внесении удобрений кущение протекает энергичнее и наступает часто на 5 дней раньше, чем без удобрений. Из работ Пауэра и др. (1963); Фоллет и Райхман (1972) известно, что при повышении содержания фосфора, доступного растениям, заметно увеличивается температурный оптимум для роста ячменя.
Приемы агротехники, обеспечивающие быстрый рост растений и создание ими густого покрова растительной массы, позволяют более полно поглощать солнечные лучи для фотосинтеза и лучше предохранять почву от нагрева. В опытах на Эрастовской опытной станции (в среднем за 2 года) при густоте 399 стеблей на 1 м2 с 27 июня по 14 июля средняя температура на поверхности почвы утром была 24,9 °С, на 1,6 °С больше, а на глубине 5 см - 21,3 °С, на 2°С больше, чем при густоте 559 стеблей. 28 июня 1958 г. в начале молочного состояния зерна в среднем за день в обычном стеблестое ячменя температура воздуха приземного слоя была 36,4 °С, а на умеренно загущенном на 8,1 °С ниже.
Изменение же среднесуточной температуры почвы на 1 - 2°С, как известно (Колясев, 1959), существенным образом сказывается на развитии корневой системы, отдельных элементов структуры урожая, соотношении репродуктивной и вегетативной части урожая, на продолжительности вегетационного периода. Обусловлено это тем, что для нормального развития в период созидательной деятельности растений температура почвы должна быть несколько ниже температуры воздуха (Радченко, 1966).
Таким образом, агротехнические приемы, способствующие развитию хорошего стеблестоя, косвенно влияют на повышение продуктивности растений в результате создания более благоприятного температурного режима. Следовательно, при совершенствовании приемов агротехники, обеспечивающих достаточное корневое питание и устойчивые запасы почвенной влаги, суховеи меньше снижают продуктивность растений ячменя.
Уровень среднесуточных температур сказывается на скорости наступления фаз развития ячменя. Чем выше температура, тем меньше продолжительность фаз, и наоборот (табл. 27). Это несомненно сказывается на росте и развитии органов, формирование которых протекает в соответствующий период, следовательно и на продуктивности растений. Для ячменя весьма благоприятны пониженные температуры во время формирования и налива зерна. Это способствует удлинению периода накопления сухих веществ и повышению крупности зерна. Опасными для растений ячменя являются высокие температуры, которые в сочетании с недостатком влаги, особенно в период формирования зерновки, вызывают снижение продуктивности растений. Так, при одинаковых условиях увлажнения в период колошение - восковая спелость (в среднем за 2 года) повышение среднесуточной температуры на 1,7 °С обусловило сокращение продолжительности фазы на 3 дня и снижение веса 1000 зерен ячменя сортов Грушевский и Донецкий 650 соответственно на 3,5 и 2,8 г.
Таблица 27. Продолжительность фаз развития растений ячменя в зависимости от среднесуточных температур воздуха (данные Эрастовской опытной станции за 15 лет)
При суховее усиливаются потери воды почвой, нарушается нормальное функционирование устьичного аппарата и повышается расход влаги на транспирацию. При недостатке почвенной влаги это вызывает преждевременное увядание вегетативной массы и резкое снижение веса 1000 зерен ячменя. В суховейном 1946 г. на Криничанском и Синельниковском госсортоучастках (Днепропетровская область) вес 1000 зерен ячменя был соответственно на 9 и 11,2 г ниже, чем в среднем за 15 лет.
Различные почвенно-климатические условия отдельных зон производства ячменя обусловливают необходимость применения дифференцированной агротехники. В северных, более увлажненных районах важное значение имеют мероприятия, направленные на улучшение условий питания растений ячменя путем применения удобрений. В засушливых районах система агротехнических мероприятий должна быть направлена в первую очередь на накопление влаги и рациональное использование ее растениями, а также на повышение эффективного плодородия почвы. Там наряду с применением удобрений пищевой режим можно улучшить за счет правильной обработки почвы.
Все подробности Ширмы настольные здесь
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'