РОСТ
РОСТ растений, биологический процесс необратимого увеличения размеров и веса р-ния и его органов. Р. р-ний обусловлен и сопровождается возникновением новых клеток, тканей, органов и веществ, из к-рых строятся структурные элементы и тело р-ния. В нек-рых случаях Р. отдельных органов сопровождается уменьшением общего веса р-ния, напр. в начальных фазах прорастания семян, клубней, луковиц, черенков, при отрастании надземных органов у многолетних р-ний после выхода их из состояния зимнего покоя. В этих случаях первоначальный Р. новых органов (корешков, стеблей, листьев) осуществляется за счёт запасов пластических веществ и сопровождается интенсивным дыханием, причём часть органических веществ окисляется до углекислоты и воды. Уменьшение общего исходного веса р-ния продолжается до образования зелёных листьев, после чего расход органических веществ на дыхание с избытком компенсируется созданием их за счёт фотосинтеза.
Рост и развитие растений - процессы нетождественные. При определённых условиях оба эти процесса могут протекать быстро; при других условиях Р. идёт быстро, а развитие медленно, или, наоборот, при быстром развитии замедляется Р. Для нормального и продуктивного Р. зелёных р-ний, в результате к-рого могут создаваться высокие урожаи, необходимо наличие в атмосфере углекислого газа, достаточное количество света, тепла и влаги, обеспеченность р-ний азотом и элементами минерального питания. Не менее важное значение имеют и другие условия внешней среды: физико-химические свойства почвы, отношение между длиной дня и ночи (фотопериоды) и т. д.
Различные проявления и сочетания этих условий по-разному влияют не только на общую интенсивность Р., но и на направление ростовых процессов, а отсюда и на структуру р-ний и урожая: на соотношение между Р. вегетативных и репродуктивных органов, на соотношение между ассимилирующими и запасающими органами (клубни, корни, луковицы и др.), на строение отдельных органов (напр., степень ветвления стеблей льна, конопли) и т. д. Характер Р. зависит и от наследственных особенностей самого р-ния, от его способности ассимилировать условия среды.
Основным элементарным процессом Р. р-ний является размножение клеток, приводящее к увеличению их числа и к формированию новых тканей и органов. Деление клеток происходит, главным образом, в особых образовательных тканях - меристемах. Для Р. в длину осевых органов р-ний особое значение имеют меристемы конусов нарастания кончиков корней (рис. 1) и верхушек стеблей (рис. 2). Это первичные ткани, т.к. клетки их произошли непосредственно от первичных клеток зародыша.
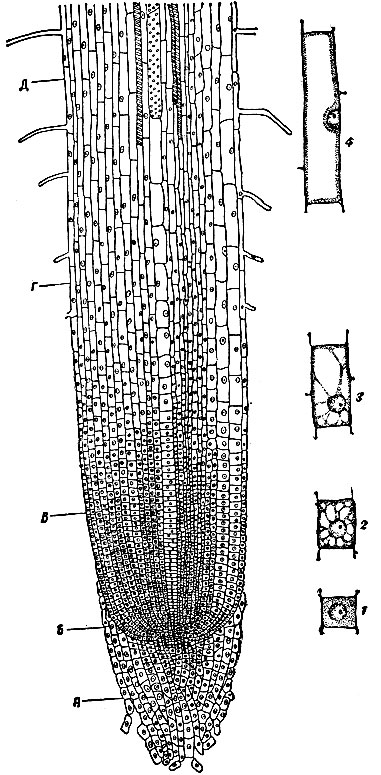
Рис. 1. Продольный разрез кончика корня бобов: А - чехлик; Б, В - зона меристемы, где происходит деление клеток; Г - зона растяжения клеток; Д - зона диференцировки клеток. 1 - 4 - клетки в разных стадиях роста
Деятельность первичных меристем, именно увеличение числа клеток, а затем растяжение и увеличение размеров вновь образованных клеток обусловливают Р. стебля в длину в верхней его части и Р. корня у его кончика. Меристемы конусов нарастания стеблей в результате деления клеток дают листовые зачатки. Конус нарастания вместе с листовыми зачатками называется верхушечной почкой. По мере возрастания числа клеток и последующего их растяжения в промежуточных участках оси побега листовые зачатки удаляются друг от друга. Так образуются междоузлия. Вся основная ткань зачатков листьев первоначально носит характер меристемы. На более поздних фазах формирования листьев в них остаются ограниченные зоны образования новых клеток: у однодольных - в основаниях листьев, у двудольных - в разных местах пластинки. Однако Р. листьев обычно прекращается довольно быстро. В пазухах листьев закладываются пазушные почки, из к-рых могут развиться боковые побеги (рис. 3).
Осевые органы растут не только в длину, но и в толщину (радиально). Это ярко выражено у стволов деревьев и у двудольных травянистых р-ний, образующих утолщенные осевые органы (напр., корнеплоды). В радиальном Р. осевых органов двудольных р-ний важное значение имеет вторичная меристема - камбий, состоящий из цилиндрического слоя деятельных клеток. Клетки камбия откладывают внутрь элементы древесины, а к периферии элементы коры. Вслед за образованием новых клеток начинается их растяжение, т. е. увеличение размеров. Переход от эмбрионального Р. к растяжению можно наглядно наблюдать на кончиках корней. Клетки, образуемые вторичными меристемами и, в частности, камбием, также увеличиваются в размерах очень сильно, иногда в диам. в 5 - 40 раз по сравнению с исходными размерами, а следовательно, в объёме в 100 - 1000 раз. Иногда различия в размерах одних и тех же органов разных р-ний зависят в большей мере от различий в размерах их клеток, чем от различий в интенсивности деятельности меристем, а следовательно, от различий в числе клеток (напр., плоды разных сортов одного вида). Культурные р-ния, образующие более крупные органы, отличаются от своих диких родичей не только более активной деятельностью меристем, но и более крупными клетками, к-рые у культурных р-ний достигают иногда громадных размеров (напр. клетки мякоти плодов арбуза, апельсинов, лимонов и мандаринов, клетки запасающей паренхимы корнеплодов и картофеля).
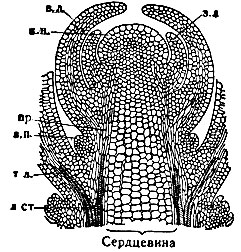
Рис. 2. Продольный разрез через верхушку стебля с точкой роста: к. к. - конус нарастания, прикрытый зачатками листьев; з. д., з. п. - зачатки пазушных почек; пр. - прокамбий; т. л. - спиральные трахеиды и сосуды развивающегося листа; л. ст. - примыкание листового пучка к лубу и древесине стебля
В общий процесс Р. р-ний входит также диференцировка клеток, в результате к-рой новые клетки специализируются, образуют ткани и выполняют различные функции (клетки и ткани проводящие, запасающие и т. д.).
Основные этапы Р. (эмбриональный, растяжения и диференцировки) у разных р-ний различны. Эти различия обусловлены прежде всего наследственными особенностями р-ний: в результате Р. образуются ткани, органы и структуры, свойственные данному виду и форме р-ния. Ростовые процессы подчиняются также многим общим для всех р-ний закономерностям, напр. строго определённая ориентировка направления Р. органов и постоянное размещение органов по отношению друг к другу.
Способность р-ний ориентировать направление Р. отдельных органов в зависимости от действия или локализации нек-рых условий внешней среды носит название тропизмов. Так, корни обладают положительным геотропизмом и растут по направлению действия силы тяжести. Стеблям же свойствен отрицательный геотропизм. Направление органов в зависимости от действия света носит название фототропизма (у корней отрицательный, у стеблей положительный), в зависимости от различной степени увлажнения - гидротропизма, а в зависимости от концентрации питательных или активных хим. веществ - хемотропизма. На направление Р. отдельных органов (особенно корней) влияют и условия аэрации. Разные органы различно реагируют на воздействие внешней среды. В конечном итоге направление Р. и положение органов зависят от неск. импульсов, действующих иногда и противоположно один другому. Так, не всегда и не все корни растут строго вертикально вниз; в зависимости от условий влажности, аэрации и локализации питательных веществ они либо глубоко проникают в землю, либо располагаются в поверхностных слоях почвы. Стеблевые органы не всегда растут строго вертикально. В зависимости от сочетания действия силы тяжести, света, связанного, напр., с густотой стояния р-ний, формируются и вырастают р-ния либо прямые, маловетвистые, высокие (как, напр., сосны в бору или лён и конопля в густом посеве), либо более низкие, ветвистые, раскидистые, как, напр., одиночные деревья или лён и конопля в изреженных посевах.
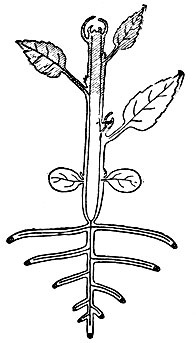
Рис. 3. Схема размещения меристем у двудольного растения. Образовательная ткань заштрихована чёрным
На ход отдельных элементарных процессов, из к-рых складывается общий Р. р-ния, одни и те же условия часто влияют по-разному. Так, напр., деление клеток в образовательных тканях конусов нарастания корешков, в почках и в камбии многих р-ний может происходить не только при наиболее благоприятных для большинства биологических процессов темп-pax в 20 - 25°, но и при темп-pax, близких к нулю. Гораздо сильнее зависит от внешних условий растяжение клеток и увеличение их размеров. Р. клеток в значительной степени определяется энергией поступления в них воды, их тургором. Недостаток воды ведёт к ослаблению и потере тургора клеток, сильно задерживает и даже приостанавливает Р. р-ний и их органов. При недостаточном водоснабжении р-ния получаются вообще мелкоклеточными (ксероморфная структура). Сильно влияют на растяжение клеток также условия освещения. В отсутствие света р-ния (напр., ростки клубней картофеля, растущие в темноте) сильно вытягиваются и несут на себе только зачаточные недоразвитые листья. Свет сильно сокращает растяжение стеблевых клеток, но способствует усиленному разрастанию листьев. При надлежащей интенсивности света р-ния не вытягиваются, вырастают крепкими, хорошо облиственными, но имеют более мелкие клетки, чем р-ния, растущие при ослабленном освещении.
Обильное питание р-ний при хорошем увлажнении способствует усиленному Р. клеток, и при высокой агротехнике р-ния имеют ткани с крупными клетками. Именно поэтому возделываемые р-ния в условиях хорошего питания и водоснабжения отличаются от соответствующих диких форм наличием крупных клеток и большими размерами органов, что связано также и с большей активностью их меристем.
В силу того, что изменение условий внешней среды по-разному влияет на Р. отдельных органов, у р-ний, выращиваемых в разных условиях, меняются соотношения между размерами различных органов. Так, напр., в условиях высокого увлажнения и ослабленного освещения относительно слабо развивается корневая система. При недостаточном водоснабжении и избыточном свете соотношение изменяется в пользу корней. Условия внешней среды оказывают сильное влияние и на ход диференцировок, что влияет на анатомо-морфологические структуры и признаки. На ход диференцировок и направление ростовых процессов сильно влияет соотношение между длиной дня и ночи (см. Фотопериодизм).
Важной особенностью Р. р-ний является ритм ростовых процессов. Ритмичность наиболее резко выражена у многолетних р-ний в соответствии со сменой условий в течение года. В неблагоприятные для жизнедеятельности периоды, напр. в течение холодной зимы или во время сухих периодов, Р. р-ний прекращается, и они переходят в состояние покоя, к-рый бывает глубоким и вынужденным. При глубоком покое в клетках р-ний происходят столь сильные качественные изменения, что р-ние, даже при помещении его в благоприятные условия, не способно к росту.
При вынужденном покое р-ния или их органы не теряют готовности к Р., но не растут, вследствие отсутствия подходящих условий (напр., низкие темп-ры, недостаток влаги и др.). Состояние покоя у р-ний или их органов (клубни картофеля, семена) нередко связано с тем, что почки и ткани, содержащие запасные вещества, защищены внешними оболочками и тканями от доступа к ним кислорода, чем задерживается нужный для прорастания обмен веществ. Обеспечивая доступ кислорода внутрь покоящихся органов (напр., очищая клубни от пробкового слоя или повреждая кожуру семян), можно обеспечить их прорастание. Регулировать состояние покоя можно и нек-рыми физиологически активными веществами. Так, этиленхлоргидрин побуждает к прорастанию покоящиеся свежеубранные клубни картофеля. Наоборот, уретаны, а также эфиры а-нафтилуксусной кислоты задерживают прорастание клубней, закончивших период естественного покоя. Обрабатывая этими веществами клубни в хранилищах, можно предохранить их от прорастания и значительно сократить происходящие от этого потери. Прекращение состояния покоя связано с набуханием клеток меристемы конусов нарастания, зародышей или вегетативных почек. После этого начинается усиленное деление клеток, а вслед за этим растяжение и диференцировка, обеспечивающие Р. осевых органов в длину. Интенсивность этого типа Р. идёт в начале вегетации в нарастающем темпе, но вскоре постепенно затухает. Однако у некоторых р-ний в течение вегетационного периода наблюдается неск. периодов усиленного Р. осевых органов (напр., у лимонов).
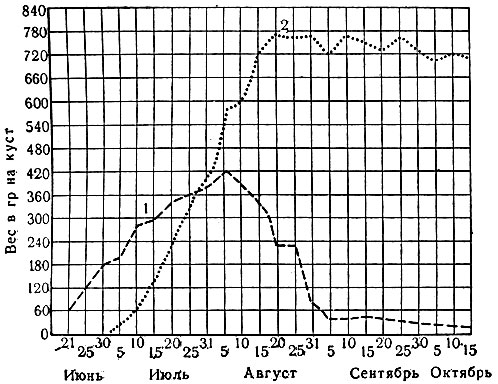
Рис. 4. Динамика накопления веса ботвы 1 и клубней 2 картофеля (по Лорху)
Деятельность вторичных меристем обычно начинается позднее и продолжается дольше. Если р-ния образуют осевые органы с сильно развитыми запасающими тканями, то вслед за усиленной деятельностью вторичных меристем начинается дальнейший усиленный вторичный Р. этих органов и тканей. Этот вторичный Р. связан с увеличением размеров ранее образованных камбием паренхимных клеток. Одновременно эти клетки заполняются запасными веществами, что предшествует завершению сезонного цикла роста.
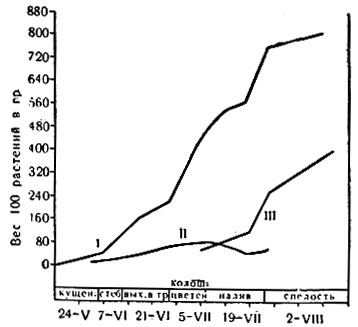
Рис. 5. Ход накопления сухого вещества пшеницы: I - всего растения; II - листьев; III - колосьев (из работы Васиной)
В процессе Р. р-ния сначала образуют органы, необходимые для питания, а затем формируют органы и ткани, служащие для размножения и для поддержания и сохранения жизни в неблагоприятные периоды. В интенсивности Р. различных органов наблюдается закономерная последовательность, что обычно связано с экологическим типом р-ния и с условиями его жизни (рис. 4 и 5).
В течение вегетации направление и характер ростовых процессов меняются несколько раз. Для наилучшего Р. разным р-ниям нужны различные условия. Умение управлять Р. в нужном направлении в каждой фазе жизненного цикла р-ний является существенным условием получения высоких и устойчивых урожаев.
См. также Вегетационный период.
Литература: Васина А., Динамика роста и развития яровой пшеницы при орошении, в сб. "Вопросы ирригации", вып. 3, Саратов, 1936 (Труды Всесоюзного института зернового хозяйства, т. VII); Комаров В., Введение в ботанику, М., 1949; Лорх А., Динамика накопления урожая картофеля, М., 1948; Лысенко Т., Агробиология, [6 изд.], М., 1952; Максимов Я., Краткий курс физиологии растений, 8 изд., М., 1948; Раздорский В., Анатомия растений, М., 1949; Ракитин Ю., Применение ростовых веществ в растениеводстве, М., 1947; Турецкая Р., Приемы ускоренного размножения растений путем черенкования, М.-Л., 1949.
Источники:
- Сельскохозяйственная энциклопедия. Т. 4 (П - С)/ Ред. коллегия: П. П. Лобанов (глав ред) [и др.]. Издание третье, переработанное - М., Государственное издательство сельскохозяйственной литературы, М. 1955, с. 670
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'