ОПЛОДОТВОРЕНИЕ У РАСТЕНИЙ
ОПЛОДОТВОРЕНИЕ У РАСТЕНИЙ, центральный акт полового процесса, заключающийся в слиянии мужской и женской половых клеток (гамет) и следующей затем взаимной ассимиляции их. Новая клетка - зигота, возникающая в результате оплодотворения, даёт начало новой жизни, из неё развивается новый организм. Оплодотворение известно у всех групп р-ний, кроме бактерий и синезелёных водорослей, к-рые размножаются бесполым путём.
Долгое время считали, что р-ния, в отличие от ж-ных, не имеют пола. Однако практика шла значительно впереди теории, и ещё в древности применялось искусственное опыление финиковых пальм для усиления плодоношения.
Впервые убедительное доказательство участия мужского и женского начала в формировании нового растительного организма дал акад. Российской академии наук Кельрейтер (1761). Он перенёс пыльцу одного вида табака на рыльце другого вида и получил гибридное р-ние, соединяющее в себе особенности обеих родительских форм. Так. обр., Кельрейтером было установлено, что у высшей группы р-ний - у покрытосеменных - пыльники, дающие пыльцу являются мужскими органами, а пестик, содержащий семяпочки, развивающиеся в дальнейшем в семена, - женским половым органом. Кельрейтер полагал, что новый организм возникает в результате слияния мужского и женского начала, но морфология этого процесса и подробности строения половых элементов ему были неизвестны. Он первый наблюдал начальные стадии прорастания пыльцы, но из своих наблюдений заключил, что мужским началом является пигмент, сползающий с пылинок в воде, а женским началом - женская семенная жидкость, происхождение к-рой он не мог выяснить.
Дальнейшее изучение полового процесса шло не без зигзагов. Так, наблюдения над прорастанием пыльцевых трубок в столбике и в завязи снова привели нек-рых учёных к теории, отрицавшей половые явления у р-ний. Такие учёные предполагали, что зародыш, уже будто бы имеющийся в кончике пыльцевой трубки, только вносится в семяпочку. В середине 19 в. было сделано несколько важных открытий. Польский учёный Лещик-Суминский (1848) открыл половой процесс у папоротников и установил, что этот процесс происходит не на олиственном р-нии, приносящем лишь споры, а на маленьком, не расчленённом на органы растеньице - заростке,- вырастающем из споры папоротника. Он показал также, что оплодотворение совершается подвижными в воде "нитями" - сперматозоидами папоротника. Гофмейстер (1847 - 1851) описал типичное строение зародышевого мешка покрытосеменных, содержащего яйцевой аппарат (яйцеклетку и две вспомогательные клетки) у одного полюса, группу из трёх клеток - антипод у другого полюса и центр. клетку с двумя ядрами, названными полярными (рис. 1). В противоречие с теорией внесения зародыша в семяпочку, Гофмейстер установил, что зародыш развивается из яйцеклетки в результате полового процесса. Гофмейстер сделал шаг вперёд в изучении полового процесса, но всё же считал, что пыльцевая трубка воздействует на яйцеклетку веществами, проходящими осмотически через её стенку, не проникая в зародышевый мешок. Так. обр., центральный акт полового процесса у семенных р-ний продолжал оставаться невыясненным. В этот же период было открыто и экспериментально доказано оплодотворение у бурых водорослей (Тюре, 1853). Этот процесс, как и развитие зиготы, происходит у водорослей в воде и доступен непосредственному наблюдению на живом материале.
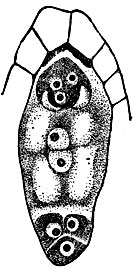
Рис. 1. Строение зародышевого мешка у мышехвостника (по Черноярову)
В последней четверти 19 в. при изучении пыльцы и пыльцевых трубок были выяснены замечательные явления, связанные с процессом оплодотворения у семенных р-ний. Эти исследования представляли большие трудности в связи с тем, что объекты изучения - клетки пыльцы и пыльцевой трубки очень мелки, значительно мельче клеток зародышевого мешка. Выдающееся значение в этих исследованиях имели труды русских учёных И. Н. Горожанкина, В. И. Беляева и С. Г. Навашина. Исследования делений, происходящих в пылинке и пыльцевой трубке семенных р-ний, показали, что пыльцевая трубка содержит оформленные элементы (клетки, ядра), к-рые во время роста пыльцевой трубки перемещаются в её кончик. Удалось также наблюдать в цитоплазме яйцеклетки сливающееся с её ядром мужское ядро. Однако немецкий цитолог Страсбургер продолжал поддерживать диффузную теорию оплодотворения. Он полагал, что оформленные элементы в кончике пыльцевой трубки расплываются, их вещество диффундирует через тонкую оболочку пыльцевой трубки и оформляется вновь в цитоплазме яйцеклетки. Ошибочность этих положений была доказана И. Н. Горожанкиным. В докторской диссертации "О корпускулах и половом процессе у голосеменных" (1880) он писал, что никакого растворения и новообразования ядер не происходит и предположил, что мужские гаметы из пыльцевой трубки проникают в яйцеклетку и сливаются с ней. В 1883 при исследовании опылённых семяпочек сосны Pinus pumilio он окончательно доказал правильность своей точки зрения. Он получил препарат, где мужская гамета была застигнута в момент прохождения через отверстие в оболочке пыльцевой трубки. Это исследование было настолько убедительным, что уже в 1884 Страсбургер опубликовал большую работу об оплодотворении у семенных р-ний, в к-рой отказался от прежней теории диффузного оплодотворения и принял непосредственный переход оформленных спермиев из пыльцевой трубки к яйцеклетке.
Работы Беляева пролили свет на эволюцию семенных р-ний. Он подробно выяснил гомологию (сходство в развитии) мужского заростка разноспоровых папоротникообразных и пыльцы и пыльцевой трубки голосеменных. Он установил, что пыльцевая трубка голосеменных показывает упрощение в строении и позволяет понять переход к пыльцевым трубкам покрытосеменных. В этих исследованиях Беляев завершил работу, начатую П. Ф. Горяниновым(1834), приведшим в эволюционную связь папоротникообразные и семенные растения.
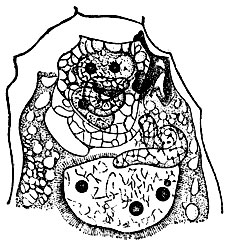
Рис. 2. Двойное оплодотворение у рябчика (Fritillaria) ceм. лилейных (по Навашину)
В 1898 С. Г. Навашин опубликовал сообщение: "Пересмотр процессов оплодотворения у Lilium Martagon и Fritillaria tenella". Сообщение это вскрыло новые замечательные явления в оплодотворении у покрытосеменных. На объектах, к-рым уделяли большое внимание в течение предыдущего десятилетия многие учёные, в т. ч. Страсбургер и французский ботаник Гиньяр, С. Г. Навашин открыл явление двойного оплодотворения (рис. 2). Было известно, что в пыльцевой трубке покрытосеменных образуются две мужские гаметы - два спермия, как они были названы в связи с отсутствием у них органелл движения, свойственных мужским гаметам ниже организованных р-ний - сперматозоидам. Один спермий, согласно наблюдениям исследователей, оплодотворяет яйцеклетку, какова же судьба второго, "лишнего", спермия - было неизвестно. С. Г. Навашин выяснил, что второй спермий не "лишний", что он участвует в оплодотворении, сливаясь с т. н. полярными ядрами центр, клетки зародышевого мешка. Так. обр., в зародышевом мешке покрытосеменных, как оказалось, при оплодотворении происходят 2 акта слияния клеток; в результате одного из них возникает зародыш, в результате второго - особая ткань, содержащая питательные для зародыша вещества, - эндосперм. Как только было опубликовано сообщение Навашина, выяснилось, что многие учёные видели двойное оплодотворение на своих препаратах, но не поняли наблюдаемого явления, считая близость второго спермия к полярным ядрам явлением случайным. Открытие двойного оплодотворения дало толчок к дальнейшим исследованиям, и за короткое время это явление было описано у многих покрытосеменных р-ний. Одним из первых, проверивших исследование С. Г. Навашина, был Страсбургер. Он проследил явление оплодотворения на живом объекте с исключительно прозрачными семяпочками - подъельнике - и полностью подтвердил правильность замечательных наблюдений русского учёного.
Так. обр., к началу текущего столетия учение об оплодотворении достигло довольно высокого уровня. Дальнейшее развитие этого учения было задержано влиянием ложной теории вейсманизма-морганизма, порождения идеологии загнивающего капиталистического общества. Многообразное, богатое по содержанию явление оплодотворения было введено лишь к механическому соединению ядер, признанных монопольными носителями наследственности. Одним из важнейших цитологических доказательств монополии ядра в наследственности считали будто бы установленное отсутствие собственной цитоплазмы у спермиев покрытосеменных. Страсбургер, соавтор теории монополии ядра в явлениях наследственности, первый выдвинул это "доказательство". Оно было поддержано также С. Г. Навашиным. Более двух десятилетий исследования, обеднённые метафизической теорией, не внесли ничего нового в познание явлений оплодотворения Работы, показывавшие наличие цитоплазмы у мужских гамет (В. В. Финн), замалчивались. Официальная наука игнорировала гениальные труды отечественного учёного И. В. Мичурина, с горечью писавшего: "...признать, что все полученные мною новые сорта выведены без всяких научных оснований, - как бы "незаконнорожденными", - как это утверждает большинство теоретиков, в сущности, полнейших профанов в практическом деле, - было бы крайне смешно..." (Мичурин И., Соч., т. I, 1948, стр. 510).
В нашей стране зародилось и с победой социализма расцвело передовое, мичуринское учение, открывшее новую блестящую страницу в познании полового процесса у р-ний. Оно показало ложность моргановской теории и разрушило мёртвые схемы (в том числе и схему оплодотворения), державшие в плену биологическую мысль. Мичурин открыл, что оплодотворение имеет свойства основного жизненного явления; оно зависит от условий внешней среды, ему присуща характерная особенность биологического явления - избирательная способность. Вместо прежних бесплодных схем морганизма, к-рые привели к полному отказу от воздействия на половой процесс, Мичурин дал основу действенной теории, позволяющей управлять им: учение о подборе родительских пар при учёте влияния жизненных условий и исторического прошлого компонентов скрещивания, учение об отдалённой гибридизации, о вегетативном сближении, о воспитании сеянцев-гибридов, о выборе цветков для скрещивания, о приёмах опыления и др.
Т. Д. Лысенко развил дальше мичуринское учение. Он обогатил практику соц. с. х-ва новыми важными научными открытиями, основанными на биологической теории полового процесса, к к-рым относятся: учение о внутрисортовых и межсортовых скрещиваниях при свободном опылении; стадийный анализ, как основа для подбора родительских пар; вегетативная гибридизация; создание жизненного потомства путём подбора и воспитания родительских пар и др. Т. Д. Лысенко дал стройную теорию полового процесса. Многочисленными работами было установлено сходство между половыми и вегетативными гибридами и доказано, что это явления одного порядка. Изменения, возникающие в результате влияния внешней среды, а также при вегетативной и половой гибридизации, имеют общую основу - взаимодействие как между организмом и внешней средой, так и между компонентами прививки - подвоем и привоем - и компонентами скрещивания - половыми клетками. Разрушая ложную концепцию морганизма о половых клетках, как независимых от условий жизни, от тела организма, Т. Д. Лысенко показал, что половые клетки - это кусочки живого тела. Они завершают его жизненный цикл и аккумулируют в себе путь развития всех предыдущих поколений. Половые клетки обладают основным свойством живого тела, способностью к обмену веществ. Оплодотворение и есть взаимный обмен веществ сливающихся половых клеток. В этом процессе обе сливающиеся клетки перестают существовать и возникает новая клетка с обогащённой наследственностью - зигота. Она соединяет в себе возможности развития обеих родительских форм и обладает поэтому большей жизненностью, большей приспособленностью к условиям внешней среды. В свете этого учения стала понятной вредность самооплодотворения, обедняющего природу организма, суживающего его приспособительные возможности и полезность перекрёстного оплодотворения, расширяющего эти возможности.
Широчайшее обобщение, вскрывшее биологическую природу оплодотворения, показавшее всю многогранность этого явления, открыло пути к новым обобщениям, к новым способам управления этим процессом. Обеднённая схема оплодотворения, как слияния половых ядер, так прочно, казалось, установленная в конце прошлого столетия, оказалась несостоятельной, как и вейсманистское учение о спермиях - голых ядрах. Было выяснено, что спермин покрытосеменных вовсе не лишены собственной цитоплазмы: в живом состоянии они всегда представляют полные клетки. Описание спермиев-ядер было результатом некритического отношения к препаратам, изготовленным при несовершенной обработке материала. Удалось даже в зародышевом мешке, где цитологическое исследование встречалось с большими трудностями, обнаружить в отдельных случаях спермии-клетки (рис. 3) (Киевская школа эмбриологов). Этим была завершена работа Беляева, подробно исследовавшего мужские половые клетки разноспоровых папоротникообразных и голосеменных и установившего их клеточную природу.
Развивается учение об избирательности половых клеток. Мичурин показал, что молодые плодовые деревца в первый год цветения скрещиваются легче, чем в более позднем возрасте. Так. обр., избирательность половых клеток вначале покоится на более широкой основе, а затем, с течением времени она суживается. Новые наблюдения над пшеницей выяснили, что её яйцеклетки способны воспринимать пыльцу уже за два-три дня до созревания пыльников данного цветка. Если в это время опылить кастрированные колосья пыльцой ржи, то семена завязываются в таком же количестве, как и при опылении пыльцой пшеницы. Однако способность завязывать семена при опылении пыльцой ржи утрачивается с началом цветения максимум через день после его начала. Вместе с тем способность воспринимать свою пыльцу резко возрастает. Дня два эта способность держится приблизительно на том же высоком уровне, а затем быстро снижается (Д. А. Долгушин). Многочисленные опыты проращивания пыльцы в искусственных культурах также показали, что избирательная способность пыльцы формируется в течение индивидуального развития. Пылинки, высеянные сейчас же после раскрытия пыльников, легко прорастают на соответствующих искусственных средах. При хранении пыльцы количество прорастающих пылинок становится всё меньшим, и, наконец, пыльца совершенно утрачивает способность прорастать. При нанесении такой пыльцы на рыльце тем не менее образуются пыльцевые трубки и происходит оплодотворение. Однако и способность прорастать на рыльце постепенно утрачивается, хотя в нек-рых случаях она сохраняется очень длительное время (год и больше). Срок сохранения жизнеспособности пыльцы зависит от вида и от условий хранения пыльцы.
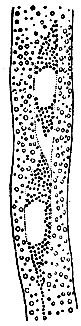
Рис. 3. Спермини-клетки у Crinum сем. амариллисовых (по Кострюковой)
Концепция моноспермного оплодотворения, требуемая моргановской теорией, не отвечает данным новых исследований. Уже давно были известны факты влияния чужой пыльцы на кожуру семени, околоплодник, плодоножку и даже прилегающие ткани материнского растения - явления так наз. метаксений. Так как эти образования принадлежат материнскому р-нию, то в метафизической схеме оплодотворения метаксениям нет места. Мичурину были хорошо известны метаксении, описанные им у гибридных р-ний. Он указывал, что половой процесс отражается и на материнском организме в его ближайших частях.
Дальнейшие исследования открыли в явлении оплодотворения новые и неожиданные стороны. Оказалось, что примесь материнской пыльцы при опылении может не исключать развития гибридного семени, но повышает жизнеспособность сеянца, вырастающего из этого семени (А. А. Авакян). Обнаружено, что примесь чужой пыльцы при искусственном самоопылении перекрёстников, если она не обусловливает развития гибридного р-ния, всё же влияет на оплодотворение, снижая депрессию и изменяя нек-рые особенности инцутируемых р-ний. Сравнение изолированного и естественного самоопыления у самоопылителей и установление депрессий при изоляции позволило понять относительно медленное вырождение самоопылителей как результат действия попадающей на рыльце чужой пыльцы (Г. А. Бабаджанян). Уже эти данные свидетельствуют о многообразии полового процесса у растений. Учение Т. Д. Лысенко об обмене веществ, как об общей основе всех жизненных явлений, в том числе и оплодотворения, открывает путь к пониманию всех этих явлений. Если рассмотреть в свете этого учения половой процесс покрытосеменных р-ний от момента попадания пылинок на рыльце материнского р-ния, окажется, что все явления представляют собой взаимодействие, обмен веществ ткалей женского органа - рыльца, столбика, завязи, семяпочки и, наконец, зародышевого мешка с пылинками, растущими из них пыльцевыми трубками и их содержимым, изливающимся в зародышевый мешок. В оплодотворении, так. обр., принимают участие не только пара спермиев одной пыльцевой трубки, но и всё её содержимое, изливающееся в зародышевый мешок и обнаруживаемое на препаратах в момент оплодотворения. Пыльцевые трубки, дополнительно попадающие в зародышевый мешок, что иногда видно на цитологических препаратах, также не являются лишними, как считали раньше,- они усваиваются развивающимся зародышем и эндоспермом. Не достигающие семяпочек пыльцевые трубки, взаимодействующие с тканями столбика и завязи, также участвуют в оплодотворении. На основе этого сложного взаимодействия идёт развитие не только яйцеклетки и центр. клетки зародышевого мешка, где оплодотворение может наблюдаться непосредственно в микроскоп но и всего пестика и даже прилегающих к нему тканей материнского растения. Так. обр., в свете этого учения находит объяснение "загадочное" явление метаксений, как и др. замечательные явления, открытые в последнее время советскими учёными. Вместе с тем исчезает метафизическое разграничение процессов опыления и оплодотворения. Оба эти явления представляются как единый процесс взаимодействия, начинающийся на рыльце р-ния и заканчивающийся в зародышевом мешке.
Однако это единое явление многообразно. И. В. Мичурин (Соч, т. I, 1948, стр. 397) учил различать в оплодотворении разные его стороны: "настоящий половой процесс", продуктом к-рого является зародыш, и "другое соединение частичек мужского и женского организма", отражающееся на материнском растении.
Новые исследования (Н. И. Фейгинсон, Н. В. Турбин, А. А. Авакян и М. Ястреб, А. Б. Саламов, Д. В. Тер-Аванесян, Э. Г. Кочарян) показали неисчерпаемость явления оплодотворения. Опыты по одновременному опылению пыльцой нескольких сортов с несомненностью установили явление участия в оплодотворении нескольких отцовских форм. Опыты были поставлены на разных р-ниях: кукурузе, томатах, пшенице, хлопчатнике - и везде получены убедительные результаты. Наблюдались случаи, когда в 1-м поколении участие 2-й отцовской формы не сказывалось, но во 2-м поколении при самоопылении появлялись р-ния с особенностями этой второй отцовской формы.
Эти наблюдения прямо ставят вопрос о полиспермии, т. е. проникновение нескольких мужских гамет в яйцеклетку. Полиспермия у ж-ных теперь цитологически доказана, доказано и внедрение живчиков в стенки матки и яйцеводов млекопитающих (Д. И. Генин). Но в отношении р-ний подобных достоверных цитологических данных ещё нет. Однако в свете мичуринской теории влияние неск. отцов м. б. понято, как и при вегетативной гибридизации, без проявления его в форме непосредственно наблюдаемого слияния клеток. Этот вопрос ждёт своего цитологического разрешения.
Учение об оплодотворении у р-ний продолжает быстро развиваться. Оно имеет твёрдую теоретическую основу в мичуринской концепции этого процесса, как жизненного явления, как взаимного обмена веществ сливающихся половых клеток. Познание этого жизненного процесса в его величайшем многообразии позволяет всё увереннее итти по пути управления им.
Литература: Арнольди В., Значение работ русских морфологов в истории науки о клетке, "Классики естествознания", кн. XII - Русские классики морфологии растений, М.-П., 1923, стр. 130 - 35; Бабаджанян Г., Заметки о явлениях полового ментора у растений, "Известия Академии наук СССР", серия биологическая, М.-Л., 1949, № 4, стр. 455 - 69; Кострюкова К., Существуют ли бесплазменные спермин у высших растений, "Яровизация", М. - Одесса, 193 9, № 5-6, стр. 153 - 61; Лысенко Т., Агробиология, [6 изд.], М., 1952; О положении в биологической науке. Стенографический отчет сессии Всесоюзной академии сельскохозяйственных наук имени В. И. Ленина 31 июля - 7 августа 1948 г., М., 1948; Турбин Н. и Богданова Е., К вопросу о природе процесса оплодотворения у растений, "Известия Академии наук СССР", серия биологическая, М.-Л., 1949, № 4, стр. 432 - 54; Фейгинсон Н., Участие нескольких отцовских форм в оплодотворении кукурузы, "Агробиология", М., 1948, № 1, стр. 92 - 108; Финн В., Об оплодотворяющих элементах и половом процессе у покрытосеменных растений, "Яровизация", М. - Одесса, 1941, № 2, стр. 7 - 12.
Источники:
- Сельскохозяйственная энциклопедия. Т. 3 (Л - П)/ Ред. коллегия: П. П. Лобанов (глав ред) [и др.]. Издание третье, переработанное - М., Государственное издательство сельскохозяйственной литературы, 1953, с. 613
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'