КЛЕТКА
КЛЕТКА, основная форма развития живого вещества, свойственная животному и растительному миру. Микроскопическое исследование открывает К. как тельце с характерной, общей для всех К. диференцировкой (цитоплазма и ядро).
История развития учения о К. Ещё Роберт Гук (1657), исследуя тонкие срезы пробки, обратил внимание на ячейки, окружённые общими стенками, и дал им название К. Однако лишь в начале 19 в. стал накапливаться материал о клеточном строении тела ж-ных и растений. В 1834 русский учёный Горянинов указал на клеточное строение как на основную особенность всех живых существ. Пять лет спустя Шванн, опираясь, гл. обр., на работы ботаника Шлейдена, обобщившего всё известное о растительной К., выяснил единство строения и развития ж-ных и растительных организмов. Шванн отметил значение ядра как постоянного клеточного образования и связал с ним возникновение и развитие новых К. Клеточная теория сыграла большую роль в дальнейшем развитии биологической науки. Она дала мощный толчок дальнейшему изучению как высших, так и низших организмов, привела к обнаружению клеточного строения простейших, к открытию клеточного деления, формированию знаний о К. и её органеллах. Глубокое познавательное значение клеточной теории было высоко оценено Энгельсом. "Познание взаимной связи процессов, совершающихся в природе, двинулось гигантскими шагами вперед... благодаря открытию клетки, как той единицы, из размножения и диференциации которой развивается все тело растения и животного. Это открытие не только убедило нас, что развитие и рост всех высших организмов совершаются по одному общему закону, но, показав способность клеток к изменению, оно обозначило также путь, ведущий к видовым изменениям организмов, изменениям, вследствие которых организмы могут совершать процесс развития, представляющий собой нечто большее, чем развитие только индивидуальное" (Энгельс, Людвиг Фейербах, 1948, стр. 38-39).
Однако вместе с накоплением знаний о К. организм метафизически был расчленён на части, и учёные занялись выяснением общности строения этих частей. Это привело к тому, что первоначальное (правда, наивное) учение о развитии организмов, данное основоположниками клеточной теории, было заменено идеалистическим учением об организме, как о сумме клеток, теорией об организме, как о государстве клеток. Это учение вполне соответствовало идеологии реакционного крыла бурж. учёных биологов (Вирхов). "Презрение к диалектике не остается безнаказанным", - говорит Энгельс (Диалектика природы, 1948, стр. 38). И действительно, уже с последней четверти 19 в. учение о К. оказалось в плену идеалистических метафизических теорий, связывающих хроматин ядра с измышленным "вечным и неизменным веществом наследственности". С этого времени внимание исследователей сосредоточилось, гл. обр., на ядре, и интересы их сильно сузились. К. стали рассматривать в полной метафизической разобщённости с условиями её жизни, ей отказывали в развитии. Всё сводилось лишь к повторению предшествовавших структур: К., по сложившимся представлениям, возникала от К., ядро - от ядра, хромосома - от хромосомы и т. д. и притом всё в неизменённом виде.
Мичуринская биология, разгромив ложные теории наследственности, новым светом озарила и микроскопическое строение организма. "В этой клетке,- говорит акад. Т. Д. Лысенко, - разнозначимы разные органеллы. Но нет ни одного кусочка, не подверженного развитию эволюции" (Агробиология, 1949, стр. 218). Совершенно исключительное значение для истинного понимания клетки имеют многолетние работы доктора биологических наук проф. О. Б. Лепешинской, к-рая на фактическом материале доказала, что К. могут возникать из неклеточного живого вещества. Постановлением Совета Министров СССР проф. О. Б. Лепешинской в сентябре 1950 присуждена Сталинская премия 1-й степени за 1949 год - за выдающиеся научные исследования неклеточных форм жизни и происхождения клеток, изложенные в научном труде "Происхождение клеток из живого вещества и роль живого вещества в организме".
Большое влияние на развитие науки о К. - цитологии оказал акад. С. Г. Навашин (1857 - 1930), имя к-рого связано с важными открытиями в области цитологии и эмбриологии р-ний (двойное оплодотворение, халацогамия, т. е. врастание пыльцевой трубки через основание зародышевого мешка, тонкое строение хромосом). Из школы С. Г. Навашина вышел ряд цитологов, содействовавших дальнейшему развитию учения о клетке: В. В. Финн (открытие спермиев-клеток у высших групп покрытосеменных, 1925), М. В. Чернояров (открытие возникновения и развития хромосом в редукционном делении, в противоречие с общепризнанным положением об их метафизической непрерывности, 1927) и др. Школой Навашина в Киеве проведено большое количество исследований живого материала, не изменённого фиксирующими жидкостями и окраской, что способствовало дальнейшему углублению знаний о клетке.
Строение К. Во всех растительных и животных К. мы находим цитоплазму (старый термин протоплазма, под к-рой теперь понимают недиференцированное, как в тельцах бактерий, или диференцированное на цитоплазму и ядро, как в К., живое вещество) и ядро (рис. 1), связанные взаимодействием в обмене веществ. С диференцировкой (поляризацией) К. на указанные образования связано возникновение самой К. Цитоплазма представляется при наблюдении на живом прозрачной и бесцветной. Во многих К. можно наблюдать течения цитоплазмы, что свидетельствует о невысокой её вязкости. Однако вязкость может значительно изменяться в процессе жизнедеятельности К. Цитоплазма разных К. отличается по своей вязкости, лучепреломляемости, а также и др. особенностям. Дискуссия о зернистом, волокнистом или пенистом строении цитоплазмы имеет сейчас лишь исторический интерес, т. к. известно, что в разные моменты жизнедеятельности цитоплазма имеет различное строение. Ядро - также бесцветное и прозрачное обычно круглое тело, содержащее в стадии "покоя" более интенсивно преломляющее свет тельце - ядрышко (рис. 2). Ядро претерпевает сложные превращения во время деления. В живом состоянии ядро м. б. обнаружено не всегда, всё же в нек-рых клетках ж-ных и р-ний оно бывает хорошо заметным, благодаря более интенсивной лучепреломляемости. В цитоплазме мы различаем так наз. клеточные органеллы, внутриклеточные живые тельца, возникающие в процессе развития клеток организма. Большое значение нек-рых органелл в дальнейшей жизнедеятельности К. (напр., пластид у р-ний) хорошо известно. Ранние исследователи К. придавали первенствующее значение оболочке. Дальнейшие исследования привели к убеждению, что оболочка имеется лишь у растительных клеток. В наст. время не подлежит сомнению, что пограничный с внешней средой слой цитоплазмы К., играющий большую роль в обмене веществ, изменён этим обменом веществ у всех решительно К. У подавляющего большинства растительных К. этот поверхностный слой уплотнён продуктами жизнедеятельности К. (напр., целлюлоза у зелёных р-ний) и образует плотную твёрдую двуконтурную оболочку. Несмотря на видимую отграниченность цитоплазмы от оболочки, связь их хорошо обнаруживается при плазмолизе (сжатии тела К. при высыхании, воздействии концентрированных растворов сахара и др.) в виде цитоплазменных нитей, идущих от твёрдой несжимающейся оболочки к отрывающемуся внутреннему слою. У животных и немногих растительных К. оболочку не удаётся непосредственно наблюдать, и эти К. представляются нам голыми. Однако в последнее время есть указания сов. ученых, что у этих так наз. "голых" К. в ряде случаев можно обнаружить при непосредственном наблюдении на живом хорошо выраженную оболочку (напр., у генеративной клетки покрытосеменных р-ний). В др. случаях эта оболочка обнаруживается при помощи особой обработки (напр., оболочка эритроцитов ж-ных). Наличие хорошо выраженной твёрдой оболочки составляет одно из морфологических отличий растительной К. от животной. Для растительных К., кроме того, характерно присутствие вакуолей - внутриклеточных полостей, выполненных клеточным соком. В клеточном соке находятся в растворённом состоянии многие вещества, возникающие в процессе обмена веществ К. р-ний: сахара, органические кислоты и их соли, алкалоиды, глюкозиды, нек-рые пигменты, дубильные вещества и т. д. Клеточный сок оказывает значительное давление на цитоплазму и упругую оболочку К., к-рая вследствие этого находится в живых растительных К. в состоянии напряжения, так наз. тургора, играющего большую роль в жизни растений. Др. различия между растительными и животными К. касаются клеточных органелл. Так, напр., особо диференцированное тельце, называемое клеточным центром, или центрозомой, м. б. обнаружено в животных К., но отсутствует в большинстве растительных К. Наоборот, только в растительных К. можно найти пластиды (рис. 3) - оформленные внутриклеточные тельца, с к-рыми связан процесс фотосинтеза (в содержащих хлорофилл пластидах - хлоропластах), а также процесс образования запасных углеводов (отложение крахмала в бесцветных пластидах - лейкопластах) и т. д. Однако у грибов и синезелёных водорослей пластиды отсутствуют. В теле высших р-ний большую роль играют отмершие К., напр. в древесине, пробке. Следует также отметить так наз. хондриосомы (рис. 2 и 3), находимые и в животных и в растительных К., и сетчатый аппарат Гольджи (рис. 4), присутствующий в животных К. и имеющий обычно вид клубка извитых канальцев. Однако данные об аппарате Гольджи получены на фиксированном материале, и картины, наблюдаемые в живых К., не соответствуют полученным при фиксировании. Высказывается мнение, что аппарат Гольджи представляет собой своеобразный вакуом (совокупность вакуолей). Образования, ранее описанные в растительной К. как аппарат Гольджи и имеющие сходное с ним строение, расшифрованы теперь как совокупность вакуолей. В К., помимо цитоплазменных образований, наблюдаются разнообразные включения, продукты жизнедеятельности, запасные питательные вещества и др. (жировые капли, капли секретов, кристаллы и т. д.).
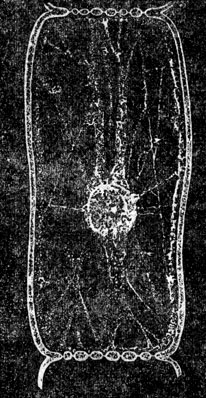
Рис. 1. Общий вид растительных клеток
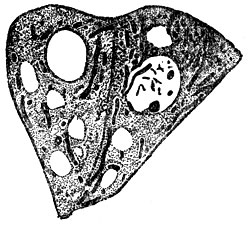
Рис. 2. Печёночные клетки лягушки (фиксировано и окрашено). Видны ядра с ядрышками, цитоплазма и нитевидные хондриосомы
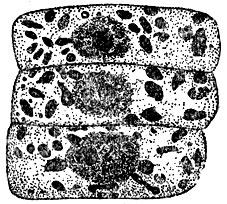
Рис. 3. Клетки листка проростка ячменя (фиксировано и окрашено). Видны пластиды - крупные и хондриосомы - мелкие зёрнышки
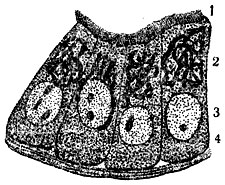
Рис. 4. Внутриклеточный сетчатый аппарат в мерцательных эпителиальных клетках: 1 - мерцательные волоски; 2 - внутриклеточный сетчатый аппарат; 3 - ядро;. 4 - цитоплазма
Как правило, К. микроскопически малы, однако величина растительных и животных К. может сильно варьировать. Нек-рые растительные К. различимы невооружённым глазом (К. мякоти плода арбуза, лимона, апельсина). У ж-ных достигают крупных размеров яйцевые К. (желток яиц птиц). Всё это нетипичные, крайние варианты. В варьировании наблюдается известная закономерность. У р-ний чаще можно встретить К. более крупных размеров, чем у ж-ных. Размеры К. в общем характерны для больших систематических групп. Так, К. беспозвоночных крупнее К. позвоночных. К. голосеменных крупнее К. покрытосеменных, однодольных - крупнее двудольных. Ср. величина К. животных колеблется от 5 до 60 - 80 μ, К. высших р-ний - 20 - 50 μ.
Форма К. также очень разнообразна и специфична для каждой ткани, отражая особенности жизнедеятельности этой ткани. Так, К. растительных волокон вытянуты, нервные К. животных обладают длинными отростками и т. д. Форма К. меняется в процессе её развития. Замечательный пример такого изменения представляет развитие генеративной К. покрытосеменных. Несмотря, однако, на указанные различия между животной и растительной К., соответствие в их морфологии (цитоплазма, ядро) и развитии не подлежит сомнению. Как указывает Энгельс, К. "...основная форма почти всякого развития жизни..." (Диалектика природы, 1948, стр. 14).
Первые исследования строения и развития К. были произведены на живом материале. Этому периоду мы обязаны нек-рыми очень тонкими наблюдениями превращений К., как, напр., описанием кариокинезиса Чистяковым (1874). Однако, начиная с конца прошлого столетия, была разработана тонкая, довольно сложная методика изготовления постоянных препаратов, к-рая позволила контрастно выявить микроскопические структуры, с трудом различимые на живом. При помощи этой методики был накоплен большой материал о строении К. Однако вместе с тем наблюдение мёртвых препаратов содействовало забвению развития К. и укреплению её понимания как застывшей мёртвой схемы. Больше того, метафизические идеалистические теории наследственности, сосредоточивая всё внимание на ядре К., вели к пренебрежению её многообразием. Под влиянием этих теорий были предложены даже такие методики, к-рые разрушали тонкую морфологическую диференцировку К., сохраняя из всего богатства клеточных структур лишь одни хромосомы. И эта разрушенная, обеднённая, до нельзя искажённая картина наивно описывалась как истинное строение К. В последнее время интерес к изучению живой К. снова возрастает. Такое изучение имеет решающее значение для понимания развития К. и в ряде случаев даёт исключительно ценный материал для выяснения тонкого её строения. Кроме того, такое изучение, несмотря на трудности, к-рые оно представляет, совершенно необходимо как надёжный контроль при изучении фиксированных препаратов. Применение ультрафиолетовой микроскопии живой К. должно способствовать дальнейшему расширению наших знаний о клетке.
Развитие К. Как и всё живое, К. представляет особую форму движения материи, не сводимую к низшим химическим, физическим и механическим движениям. "Жизнь, - говорит Энгельс, - есть способ существования белковых тел, и этот способ существования состоит по своему существу в постоянном самообновлении химических составных частей этих тел" (Анти-Дюринг, 1948, стр. 77). Основным условием самообновления является взаимодействие со средой. Как между цитоплазмой и ядром, а также др. органеллами К., так и между К. в целом и внешней средой происходит взаимодействие, обмен веществ, являющийся основой её развития. В процессе этого взаимодействия в К. возникают и исчезают включения, иногда хорошо обнаруживаемые морфологически: зёрна крахмала, капли жира, белковые и др. включения, формируются вакуоли, наполненные клеточным соком. Все эти вещества, как удаётся точно установить, по морфологическим, а также и по физ. и хим. особенностям несут печать природы К., в к-рой они возникли. Так, зёрна крахмала, столь обычные в растительных К., различаются, в зависимости от вида и характера К., как по морфологии, так и по темп-ре, при к-рой они образуют коллоидный раствор. Эти различия обмена вещества К. в различных организмах отчасти зависят от наследственной природы их, отчасти развиваются и видоизменяются в течение жизни особи в связи с разнообразными условиями взаимодействия её с окружающей средой. Так развивается целостный организм, к-рый "...не является ни простым, ни составным, как бы он ни был сложен" (Энгельс, Диалектика природы, 1948, стр. 170). Замечательные работы по вегетативной гибридизации вскрывают, как глубокий тонки различия в обмене веществ организмов, и вооружают человека совершенными способами воздействия на организмы, управления их развитием.
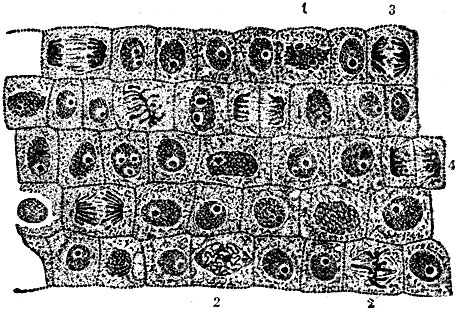
Рис. 5. Группа клеток из меристемы эмбриональной ткани лука (продольный срез растущего кончика корня): 1 - профаза; 2 - метафаза; 3 - анафаза; 4 - ранняя телофаза
Микроскопические наблюдения открывают в жизнедеятельности К. два периода: период так наз. "покоя", когда внутренняя форма К. остаётся в основном без изменений, и период деления - период глубоких превращений, ведущий к формированию 2 новых К. В периоде покоя идёт деятельный обмен веществ К. с окружающей её внешней средой, с другими К. и неклеточными структурами организма. Однако, хотя морфологические превращения и незаметны в "покоящейся" К., в течение этой стадии накопляются изменения, к-рые делают продолжение развития в той же форме невозможным, и тогда наступает деление К. Известны два способа деления клетки: митоз, или кариокинезис, и амитоз. Кариокинезис характеризуется сложными быстро протекающими изменениями ядра, хорошо наблюдаемыми на фиксированных и окрашенных препаратах (рис. 5). Хроматин - красящееся вещество ядра - образует вначале тонкую нить, свёрнутую клубком. Эта нить делится на отдельные отрезки - хромосомы, затем исчезает оболочка ядра (стадия клубка или профаза). Хромосомы располагаются в экваториальной плоскости К., образуя правильную фигуру. Каждая хромосома делится вдоль (стадия метафазы). Дальнейшее расхождение дочерних хромосом к полюсам (стадия анафазы) ведёт к образованию дочерних клубков и делению тела К. (стадия телофазы). В амитозе деление ядра происходит путём перешнуровывания иногда на неравные части. Метафизическое мышление, видевшее в митозе "механизм" для точного разделения хроматина - носителя пресловутого "вещества наследственности", придавало исключительное значение митозу. Что касается амитоза, то установилось мнение, что амитоз встречается лишь в патологических случаях. Однако уже прежние исследования, а в особенности исследования советских учёных показали, что амитоз распространён довольно широко. Обнаружен ряд переходов от амитоза к митозу в виде нетипических амитозов, нетипических кариокинезисов. Таково, напр., деление К. нек-рых простейших (амёба террикола), где фигуры кариокинетического деления наблюдаются при сохранении оболочки ядра, к-рое затем перетягивается и делится, как в амитозе. Др. примером может служить деление К. эпителия яйцеводов нек-рых грызунов. В этих К. ядра делятся амитотически, но при этом наблюдается удвоение хроматиновых зёрнышек и правильное их расположение. Эти данные показывают, что между амитозом и митозом нет непереходимой границы, воздвигнутой моргановской теорией. Они свидетельствуют, что сложное деление К. не возникло сразу, а имеет свою историю. Амитоз представляется тогда, как стадия исторического развития деления К. Неправильным является и др. положение моргановской теории, рассматривающее кариокинезис как превращение лишь ядра клетки. Наблюдения на живом и особенно микрокиносъёмки живой делящейся клетки ("Деление клеток тычиночных волосков традесканции", фильм В. И. Фаворского) убедительно показывают, что деление есть превращение К. в целом, её развитие. Деление К. заканчивается формированием 2 дочерних К. Дочерние К. морфологически сходны с материнской, но они не представляют её копий, как это утверждали метафизические теории. В теории стадийного развития р-ний раскрыто значение этих превращений. Стадийные изменения совершаются в точке роста. Как известно, в точке роста происходит не только рост живой массы р-ния, выражающийся морфологически в увеличении числа К., но и накопление незаметных количественных изменений, приводящих к коренным качественным изменениям. Поэтому К., возникающие последовательно в процессе деления, не могут быть тождественными, хотя они и сходны морфологически: каждое следующее деление приближает их к завершению текущей стадии развития, к переходу на новую стадию развития. В нек-рых случаях, однако, различия между К., возникающими в последовательных делениях, м. б. морфологически хорошо выраженными, как, напр., при 1-м делении в пыльце покрытосеменных. Изучение этого деления вскрывает и др. закономерность развития: так наз. дочерние К. (вегетативная и генеративная) значительно отличаются одна от др. В то время как вегетативная К. сходна с материнской - первичной К. пыльцевого зерна, генеративная значительно от неё отличается. Отсюда следует, что в кариокинетическом делении происходит раздвоение на новое и старое. По существу, в процессе деления материнская К. порождает одну отличающуюся от неё дочернюю К. Нет сомнения, что такое же раздвоение на новое и старое происходит и в обычных соматических делениях, но оно не имеет здесь своего морфологического выражения.
В процессе развития половых К. из неполовых происходят глубокие изменения обмена веществ К., проявляющиеся морфологически в сложных картинах, так наз. редукционного деления. Эти удивительные превращения привлекали внимание исследователей с конца прошлого столетия, но
истолковывались почти без исключения в духе менделевско-моргановской теории. Лишь мичуринская теория даёт правильную основу для понимания этих явлений.
Исторический процесс возникновения К. освещён указаниями Энгельса. К. возникла в отдалённую эпоху появления жизни на земле, когда создались условия для диференцирования комочков живого вещества на цитоплазму и ядро. Это был важный шаг вперёд в развитии живой природы, "...с этой первой клеткой, - говорит Энгельс, - была дана и основа для формообразования всего органического мира" (Диалектика природы, 1948, стр. 15).
Литература: Александров В., Анатомия растений, М.- Л., 1937; Бородин И., Курс анатомии растений, М.- Л., 1938; Глущенко И., Базавлук В., Медведева Г., О так называемых химерах, журн. "Агробиология", М., 1947, № 3; Жуковский П., Ботаника, 3 изд., М., 1949; Кострюкова К., К биологическому пониманию развития пыльцевого зерна, журн. "Агробиология", М., 1948, № 2; Крашенинников Ф., Лекции по анатомии растений, М.-Л., 1937; Лепешинская О., Клетка. Ее жизнь и происхождение, М., 1950; её же, Происхождение клеток из живого вещества и роль живого вещества в организме, 2 изд., М., 1950; Лысенко Т., Агробиология (Работы по вопросам генетики, селекции и семеноводства), [5 изд.], М., 1949; О положении в биологической науке. Стенографический отчет сессии Всесоюзной академии сельскохозяйственных наук им. В. И. Ленина 31 июля - 7 августа 1948 г., М., 1948; Русские классики морфологии растений, в сборнике "Классики естествознания", кн. 12, М., 1922; Чернояров М., "Hoвi факти в редукцiйому дiленнi у Nayas major All, та ix значения для хромозомних теорiй спадковости, "Bicник Киiвського ботаничного саду", Киiв, 1927, вып. 5 - 6. См. также Ботаника.
Источники:
- Сельскохозяйственная энциклопедия. Т. 2 (Ж - К)/ Ред. коллегия: П. П. Лобанов (глав ред) [и др.]. Издание третье, переработанное - М., Государственное издательство сельскохозяйственной литературы, 1951, с. 624
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'