5.2. Проявление и наследование признака нематодоустойчивости
Для успешной селекции на иммунитет исключительно важную роль играет правильный подбор исходных родительских форм на основе изучения комбинационной способности и закономерностей наследования признака, обусловливающего устойчивость к галловой нематоде.
Реакция высших растений на поражение нематодами контролируется генетическими системами, аналогичными тем, которые определяют их взаимодействие с фитопатогенными грибами (Щербаков, 1973).
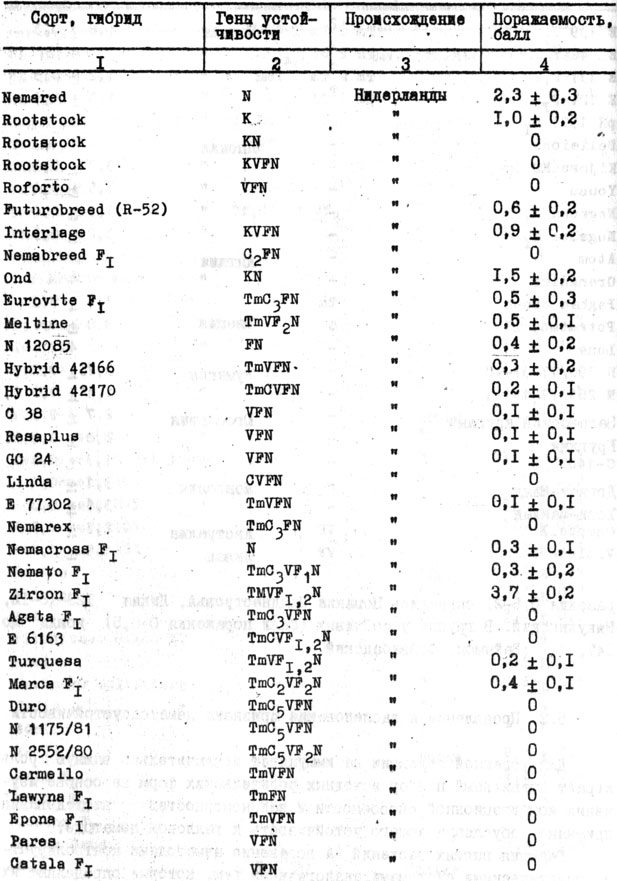
Таблица 26. Реакция сортов и гибридов томата, содержащих ген нематодоустойчивости, на поражаемость M.inoognita

Окончание таблицы 26
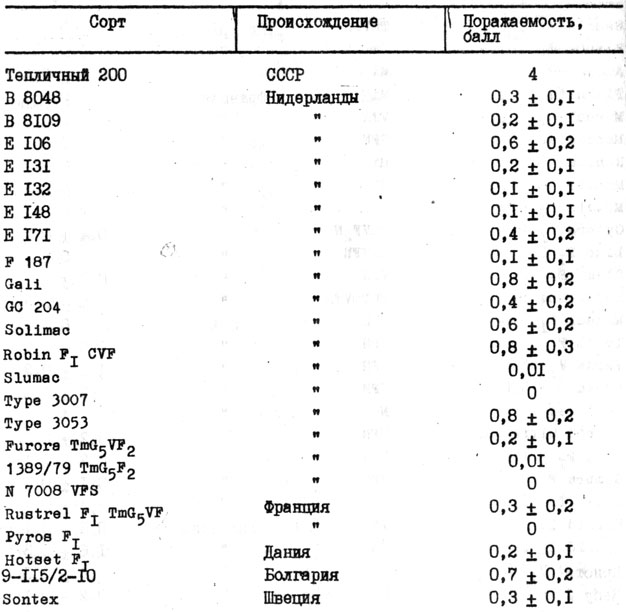
Таблица 27. Идентифицированные генетические источники устойчивости томата к галловой нематоде
Генетику устойчивости томата к галловым нематодам изучали многие исследователи. W.A. Prasier, R.K. Dennet (1949) впервые отметили, что устойчивость томата к галловым нематодам контролируется небольшим количеством доминантных генов. Затем J.C. Gilbert, d.c. Mc:Guire (1956) идентифицировали специфический способ наследования и установили присутствие единственного доминантного R-гена Mi, эффективного против всех видов Meloidogyne, за исключением M.hapla. Ген Mi находится в сцеплении в группе U. Результаты других работ свидетельствовали о единственных R-генах в сортах томата с различными типами наследования (табл. 28). Изменения в типах наследования могут происходить из-за того, что используются нечистые культуры нематод. Это подтвердилось выделением различных культур из полевых популяций с различной степенью патогенности на одних и тех же или различных сортах. Местонахождение таких культур, иногда называемых В расами, отмечали в отдельных оотеках Meloidogyne (Riggs, winatead, 1959). Это были первоначальные попытки поиска новых R-генов у томата.
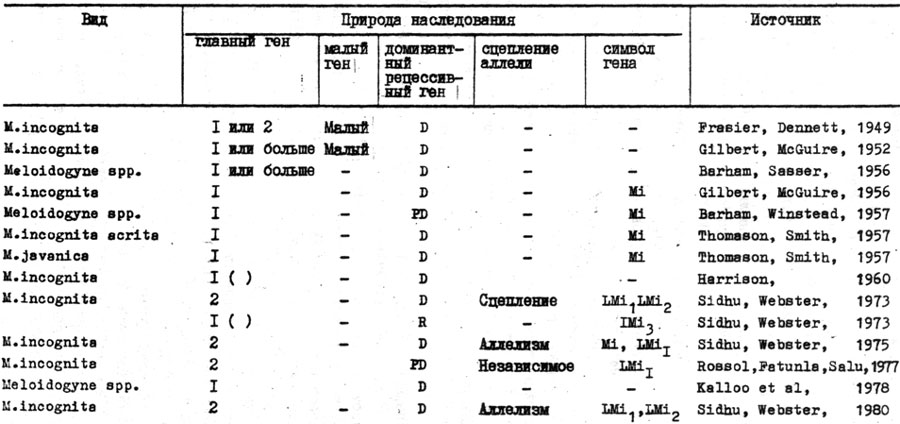
Таблица 28. Генетическая система устойчивости хозяина (L.esulentum) к галловым методам (Meloidogyne spp.) (Sidhy, Webster, 1981)
В настоящее время идентифицированы два доминантных и один, возможно, рецессивный ген и обозначены как LMiR1, LMiR2 и LmiR3 (Sidhu, Webster, 1973). Доминантные R-гены LMiR1 и LMiR2 найдены в сортах томата Nematex и small Fry-1. Третий рецессивный R-ген Lmir3, первоначально выявленный в сорте Cold Set-1, позже был признан как непостоянный в результате цитоплазматического влияния внешней среды (Sidhy, Webster, 1975). Ген LMiR1 сходен или является аллелью с локусом Mi. Принимая во внимание, что ген LmiR2 тесно сцеплен с геном LMiR1 и находится от него на расстоянии 5,65 морганид (Sidhy, Webster, 1975), T.Patunla и A.Salu (1977) идентифицировали еще один не полностью доминантный R-ген в сорте Rossol, который отличается от LMiR1, но взаимоотношение гена LMiR2 с другими R-генами не установлено. Доказана аллельность гена LMiR1 (Sidhy, Webster, 1981).
Для обозначения R-генов томата выбрана новая номенклатура (виды Meioidogyne являются полифагами). Например, старый символ Mi не указывает хозяина, поэтому добавлено L - Lycopersicon для обозначения гена видов хозяина (Sidhy, Webster, 1973). Несмотря на то, что первоначально поддерживалось добавление к записи R, указывающего устойчивость, предполагается, что его можно опустить, так как не может быть идентифицирован соответствующий ген(ы) вирулентности к видам Meioidogyne имеющимися методами генетического анализа. Кроме того, только R-гены могут быть идентифицированы в паразитических системах Meioidogyne, то есть LMi1 может быть только символом устойчивого гена.
Таким образом, большая часть результатов исследований позволяет утверждать, что устойчивость томата к галловым нематодам доминантна по отношению к восприимчивости и контролируется одним или двумя доминантными генами при возможном наличии генов-модификаторов (Rick, Butler, 1956).
Некоторые ученые (Barham, Sasser, 1956; Hare, 1965; Laterrot, 1973) считают, что доминантный ген Mi контролирует устойчивость только к таким видам галловых нематод, как южная, хлопковая,- яванская, арахисовая, но не обеспечивает устойчивости к северной нематоде.
Поражаемость нематодоустойчивых сортов vfn-8, vfn-368 галловой нематодой свидетельствует о том, что только один ген Mi не придает устойчивости ко всей популяции этого вида (Sicora, Sitara-maiah, Singh, 1973). Устойчивость сорта Nematex, имеющего ген Mi, к яванской галловой нематоде разрушается при температуре выше 28°С (Dropkin, 1969; William, Carter, 1982; Abawi, Barker, 1984). Во втором поколении гибридов наблюдается расщепление устойчивости в соотношении 3:1 (Prasier, Dennett, 1949; Kalloo, Bhatti, 1978), однако в некоторых гибридных комбинациях получали расщепление 13:3 (Gilbert, McGuire, 1952).
Таким образом, результаты приведенных исследований доказывают, что устойчивость томата к галловым нематодам определяется в основном моногенно.
Селекция томата на устойчивость к галловым нематодам достигла за рубежом больших успехов. Это направление затруднено существованием различных видов и рас галловых нематод. Основными методами селекции нематодоустойчивых сортов являются межвидовая и межсортовая гибридизация. Первые нематодоустойчивые сорта Anahu и VFN-8 получены методом межвидовой гибридизации, а сорта Ronita, Pieraita, Monita - методом межсортовой гибридизации с использованием бек-кроссов (Laterrot, 1968) устойчивого к нематоде сорта Anahu с промышленными сортами Roma, Saint Pierre, Moneymaker. Методом сложных насыщающих скрещиваний получен сорт Piersoi с комплексной устойчивостью к нематоде и некоторым заболеваниям (Жученко, 1973).
Селекционная работа по созданию нематодоустойчивых сортов и гибридов томата проводится в Нидерландах (цит. по: Снопова, 1973), Италии (Monti, Saccardo, 1977), Франции (Laterrot, 1973), Болгарии (Jordanov, stamova, 1985), Румынии (Poli, 1971), США (Lambeth, 1978), Индии (Sicora, Sitaramaiah, Singh, 1973), Иордании (Abu-Gharbien, Makkouk, Saghir, 1978), Японии (Imakawa, 1978). В СССР селекционная работа по выведению нематодоустойчивых сортов и гибридов томата для защищенного грунта ведется в НИИ овощного хозяйства (Кондакова, Игнатова, 1976), на Адлерской овощной опытной станции (Беляев, 1977; Велиев, Третьяков, 1981) и в Молдавском НИИ овощеводства (Садыкин, 1974, 1978; Садыкин, Загинайло, 1981). Как явствует из приведенного обзора литературы, сведения о генетической природе признака нематодоустойчивости у томата немногочисленны и в некоторой степени противоречивы.
Степень доминирования зависит от исходных родительских форм. Н.Н. Балашова (1977) отмечает, что доминирование, а в отдельных случаях-сверхдоминирование устойчивости у гибридов первого поколения можно объяснить более широкой амплитудой их фенотипической изменчивости по признаку устойчивости по сравнению с амплитудой фенотипической изменчивости восприимчивых компонентов. Анализ степени доминирования признака нематодоустойчивости у 200 гибридов первого поколения, полученных от скрещивания устойчивых компонентов с восприимчивыми, свидетельствует о доминантном характере устойчивости, (табл. 29); в некоторых случаях наблюдается промежуточное доминирование и сверхдоминирование (рис. 10). В тех комбинациях скрещивания, где в качестве устойчивой формы взята среднепоражаемая (балл поражения 1,6 - 2), не наблюдалось доминирования. У 25 % гибридов установлено отрицательное сверхдоминирование, у 40 - отрицательное доминирование, у 23 - промежуточное наследование, у 7 - положительное доминирование и у 5% - положительное сверхдоминирование. Высокая степень доминантности определяется минусовыми величинами, так как оценка устойчивости построена по нисходящей шкале.
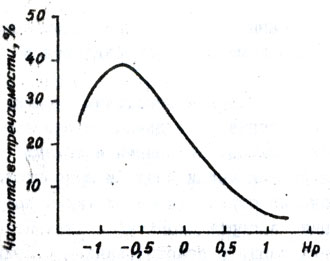
Рис. 10. Эмпирическое распределение степени доминантности нематодоустойчивости
Не выявлено влияние местоположения устойчивого родителя на нематодоустойчивость томата в гибридных комбинациях. Степень доминантностй нематодоустойчивости у гибридов первого поколения, где устойчивый сорт взят в качестве материнского, составляла 0,92±0,11, а отцовского -1,14±0,21 и варьировала в пределах ошибки опыта.
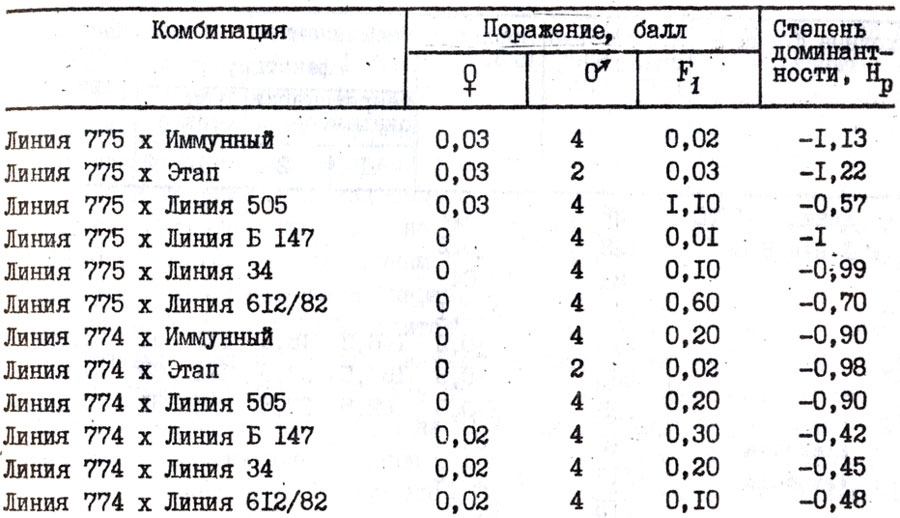
Таблица 29. Проявление нематодоустойчивости у гибридов пepвого поколения томата (1982 - 1984 гг.)
При использовании в качестве устойчивого родителя константных до отепени устойчивости линий 774/80 и 775/80 у гибридов F1 проявлялось доминирование нематодоуотойчивости (рис.11). В первом поколении гибридов, где в качестве устойчивого родителя взяты линии, отобранные из сортов Piernita, Nemared, значительно увеличивается показатель степени доминантности признака устойчивости в направлении отрицательного сверхдоминирования (до -7), из сортов Anahu, Monita в некоторых комбинациях скрещивания проявляется даже положительное доминирование (отдельные растения гибрида сильно поражаются). При использовании F1 Nemaorose отмечено значительное варьирование по степени доминантности - от +3,5 до -10.
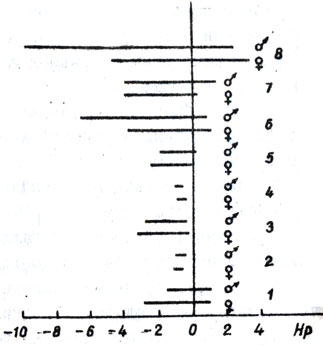
Рис. 11. Варьирование степени доминантности у гибридов томата в зависимости от местоположения устойчивого родителя: 1 - Anahu; 2 - Линия 775/80; 3 - Линия 170/70; 4 - Линия 774/80; 5 - Piernita; 6 - Линия 343/83; 7 - Линия 418/72; 8 - F1 Nemacross
Итак, у гибридов томата, полеченных от скрещивания устойчивых к галловой нематоде сортов и форм с восприимчивыми, признак нематодоустойчивости доминирует, хотя в некоторых случаях он проявляется не полностью.
Во втором поколении гибридов по характеру расщепления в основном все растения разделялись на устойчивые и восприимчивые. Расщепление согласовывалось с теоретически ожидаемым, монофакторным 3:1 (табл. 30). Таким образом, нематодоустойчивость контролируется монофакторно и с помощью беккроссов вместе с индивидуальным отбором на жестком инвазионном фоне может дать желаемый быстрый селекционный эффект по устойчивости (табл. 31).
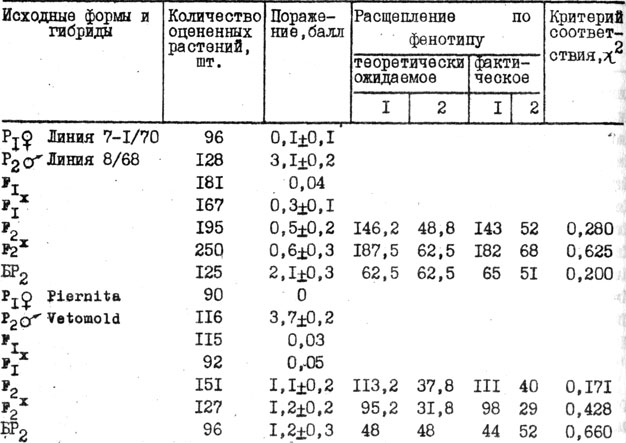
Таблица 30. Характер наследования устойчивости томата к галловой нематоде при искусственном заражении
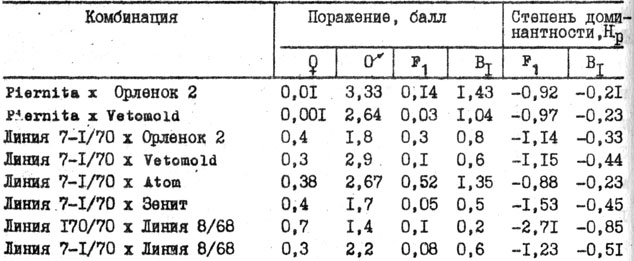
Таблица 31. Проявление нематодоустойчивости при беккроссах
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'