3.2. Разработка приемов создания инвазионного фона
В работе по созданию устойчивых к галловой нематоде сортов и гибридов томата необходимо уделять особое внимание равномерности инвазионного фона, играющего важную роль в объективной оценке селекционного материала. Это предусматривает в первую очередь изучение численности инвазионных личинок мелойдогин в тепличном грунте, где выращиваются растения томата различной степени устойчивости к фитогельминту и в зависимости от культурооборота. Вопросы методики создания инвазионного фона при селекции томата на нематодоустойчивость в литературе практически не освещены.
Количество нематод в грунте не постоянно в течение вегетации: чем старше по возрасту зараженное растение, тем больше накапливается в его корнях самок галловых нематод. Наличие инвазионных личинок и равномерность их распределения в грунте экспериментального участка можно определить путем контрольного посева семян восприимчивых к мелойдогинам растений, лучше всего огурца. На корнях последних уже через две-три недели после появления всходов можно видеть небольшие галлы. Недостатком этого метода является невозможность установления количества личинок в почве.
Степень устойчивости томата к галловой нематоде существенно влияет на количество инвазионных личинок в почве. Результаты наших исследований показали, что бессменное выращивание устойчивых и восприимчивых сортов томата создавало на участке неравномерную заселенность грунта нематодой. Степень заселенности его инвазионными личинками изменялась в течение периода вегетации и в зависимости от времени года. Меньше всего личинок насчитывали в сентябре-октябре после летнего просушивания и прогревания грунта солнцем. В сентябре после высадки растений осенне-зимнего оборота количество нематод снижалось за счет внедрения в корни поражаемых растений, а затем начинало увеличиваться и доходило до 5 тыс./экз./дм3 в январе (конец вегетации) (рис. 6). Инвазионность почвы личинками перед высадкой растений зимне-весеннего оборота составляла 2 - 5 тыс. экз./дм3. В течение января - первой половины марта количество мелойдогин уменьшалось в 2 - 3 раза как под восприимчивыми, так и под устойчивыми образцами в результате внедрения их в корни растений томата. Начиная со второй половины марта и до середины июля заселенность грунта нематодой под растениями восприимчивых форм увеличивалась в 5 - 8 раз, достигая к середине июля самой большой плотности заселения 15 - 17 тыс.экз./дм3 (табл. 12).
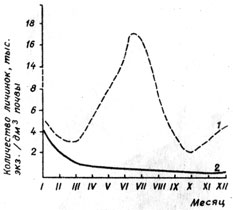
Pис. 6. Заселенность ризосферы инвазионными личинками М.incognita в динамике в зависимости от условий выращивания сортообразцов томата: 1 - восприимчивых, 2 - устойчивых

Таблица 12. Заселенность тепличного грунта галловой нематодой в зависимости от устойчивости растений (1974 - 1977 гг.)
Наблюдаемые спад и подъем численности нематоды связаны с цикличностью развития фитогельминта. Как указывает С.П. Дементьева (1971), на развитие одного поколения нематоды требуется 30 - 40 дней (в зависимости от температуры). Поэтому после высадки растений происходило снижение количества мелойдогин в грунте, а примерно через 30 - 40 дней их численность начинала увеличиваться.
Количество нематод в грунте зависело не только от продолжительности выращивания растений томата, но и от температурного режима воздуха и почвы в теплице. Так, в зимне-весеннем обороте, когда температура по сравнению с осенне-зимним оборотом выше, интенсивность развития галловой нематоды также повышалась. Растения восприимчивых образцов в конце вегетации поражались на 3,8 - 4 балла, а в осенне-зимнем обороте - на 2 - 3,3 балла.
Таким образом, инвазионность грунта на участках выращивания восприимчивых образцов в конце зимне-весеннего оборота колебалась в пределах 15 - 17, осенне-зимнего - 2-5 тыс.экз./дм3. Под устойчивыми растениями как в первом, так и во втором оборотах обнаруживали значительно меньше инвазионных личинок - 500 - 1000 и 300 - 400 экз./дм3 соответственно.
В результате многолетних исследований установлено, что нематодоустойчивые сорта и гибриды не только не снижают урожай в условиях распространения мелойдогиноза, но, что очень важно, способствуют оздоровлению грунта, его очищению от инвазии. При выращивании устойчивого к галловой нематоде сорта томата в течение вегетационного периода в теплице численность мелойдогин в почве снижается в 10 раз. Это имеет большое значение не только для защищенного грунта, но и для открытого грунта южных районов страны, где почва сильно заражена галловыми нематодами.
Как показали результаты определения заселенности грунта инвазионными личинками мелойдогин, в конце вегетации в ризосфере корней в рядках скапливалось в 2 - 3 раза больше нематод, чем в междурядьях.
Инвазионные личинки галловой нематоды мигрируют и в глубину почвенного горизонта тепличного грунта. Количество мелойдогин на различной глубине зависит также от степени устойчивости выращиваемого сорта. Так, в конце зимне-весеннего оборота под устойчивыми растениями томата при определении заселенности через каждые 5 см по вертикали не обнаружено галловой нематоды. На участке выращивания восприимчивых сортообразцов заселенность грунта верхнего слоя (0 - 5 см) личинками составляла 2600 экз./дм3, а в последующих слоях (5 - 25 см) число мелойдогин постепенно снижалось и достигало на глубине 20 - 25 см от поверхности почвы 300 экз./дм3. В более глубоких слоях (25 - 50 см) не обнаружено инвазионных личинок галловых нематод. И это вполне закономерно, так как основная масса корней растений томата (80 - 95%) находится в слое почвы до 30 см.
Зависимость развития галловой нематоды от растения-хозяина и от температуры почвы проявляется в основном в изменении сроков онтогенетического развития и половой продуктивности самок. Сроки развития нематод в корнях восприимчивых сортов томата почти в 2 раза короче, чем в корнях устойчивых. Так, время от внедрения инвазионных личинок южной галловой нематоды до выхода личинок нового поколения в корнях растений восприимчивого сорта Тепличный 200 составляло 45 дней, а в корнях устойчивого образца Линия 7-1/70 - 55 - 57 дней при средней температуре почвы 24 - 29°С, при более низкой температуре развитие замедлялось на несколько дней (Дементьева, Садыкин, 1982).
Из изложенного следует, что в устойчивых и. выносливых растениях нематода развивается медленнее, следовательно, дает меньшее число генераций за один и тот же вегетационный период. В корнях таких растений накапливается меньшее количество нематод. Этими факторами и объясняется разница в содержании мелойдогин в почве, взятой непосредственно из-под корней томата восприимчивых и устойчивых форм. Галловые нематоды как компоненты биоценоза адаптируются к его ведущим факторам, их развитие находится в прямой зависимости от биотических и абиотических факторов внешней среды. Действие этих факторов осуществляется в основном через растения, в корнях которых происходит развитие всех стадий паразита.
В связи с неравномерным характером заселения грунта нематодой для выравнивания инфекционного фона ежегодно перед высадкой селекционного материала на постоянное место под каждое растение (в лунку) вносили дополнительно по 200 г сильно инвазированной личинками почвы из расчета 13 - 15 тыс.экз./дм3. Это способствовало более равномерному распределению инвазионного начала и объективной оценке поражаемости растений. Зараженную почву вносили как под растения селекционных образцов, так и под стандартный сорт Тепличный 200, восприимчивый к галловой нематоде, который размещали через 8 - 10 рядков оцениваемых образцов.
Таким образом, заселенность почвы теплицы инвазионными личинками галловой нематоды изменялась от 2 - 5 до 15 - 17 тыс. на участках выращивания восприимчивых сортов и от 0,3 - 0,4 до 0,5 - 1 тыс.экз./дм3 - под растениями устойчивых образцов в зависимости от оборота. В конце зимне-весеннего оборота количество нематоды под восприимчивыми растениями увеличилось в 10 раз, по сравнению с устойчивыми.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://agrolib.ru/ 'Библиотека по агрономии'